|
Introducción:
Como un profesor de zoología de
invertebrados, una de las preguntas que le digo a mis alumnos
que deben responder para cada animal que examinan es "¿cómo
se alimenta?" La alimentación es el motivador
principal para los animales. Aún cuando algunos animales
pueden estar sin alimento por algún tiempo debido a
comportamientos relacionados con la reproducción, si
el animal no se puede alimentar, todo lo demás no importa;
sin comida morirá. Las diferencias de la morfología
en el aparato para la alimentación son algunas de las
formas básicas en las que los animales difieren y se
reflejan en las variadas formas corporales que los animales
adquieren.
La gente está acostumbrada a pensar
en los animales como criaturas móviles, moviéndose
de un lugar a otro. Generalmente, cuando piensan en estos
movimientos, la mayoría de la gente nota que son debidos
a la necesidad de comida del animal. Los herbívoros
se mueven cuando acaban con las plantas comestibles en cualquier
lugar y los depredadores se mueven siguiendo a sus presas.
Pero, ¿qué hay con los animales que no tienen
que perseguir su comida? ¿cómo se manifiesta
la "necesidad de comer" en cambios en su comportamiento
o morfología? A menudo los cambios se observan en otros
aspectos de su historia natural. Muchos buenos ejemplos se
hallan en los animales que se alimentan por suspensión
en los ambientes marinos de aguas someras como los arrecifes
de coral.
Para los animales que viven en los ambientes
marinos someros, la comida casi nunca es escaza (un hecho
que es muy importante en el cuidado de animales de arrecife
y que se ha comentado numerosas ocasiones en las páginas
de esta revista- vean los artículos previos de Eric
Borneman 1,
2,
3,
4,
5,
6,
7
y el Mío).
La cantidad de vida, de todas las formas y tamaños,
en el ambiente plancktónico, está más
allá de la facil comprensión. Adicionalmente,
las aguas marinas someras están siempre en movimiento,
así que toda esta vida está moviéndose
constantemente y podría estarse moviendo también
por voluntad propia.
Esto significa que el agua en los ambientes
marinos someros, hasta una profundidad de unas cuantas decenas
de metros, está llena de pequeños organismos
vivos. Estos pequeños organismos vivos son siempre
alimento para algún otro organismo, así que
básicamente el agua en esta capa del océano
es un bar de bocadillos móviles esperándo ser
colectados. Para un animal que puede utilizar algún
método para obtener la comida del agua, el alimento
nunca escacea; cada ola, corriente o flujo trae una nueva
dotación de golosinas para ser consumidas.
No es sorprendente entonces, que la selección
natural haya favorecido el desarrollo de muchas formas de
colectar esta comida. Hay varios tipos de animales que se
alimentan en suspensión pasivos o activos, así
como animales que podrían considerarse como depredadores
más activos de componentes específicos de las
comidas en el agua. Tales animales por lo general no son muy
selectivos con la comida. Si flota o nada, tratarán
de comérselo. Probablemente debido a esta no selección
generalizada, los aficionados a menudo encuentran que estos
animales son faciles de mantener con vida, al menos por un
tiempo. Si alguna comida relativamente nutritiva se coloca
en el agua, los animales se la comerán.
Consecuentemente, la mayoría de
los animales que los aficionados al arrecife mantienen caen
dentro de la categoría de animales que se alimentan
directa o indirectamente de planckton. Sin embargo, lo que
estos animales de alimentacín por suspensión
no hacen, es competir por la comida. Normalmente, para este
tipo de animales siempre hay suficiente comida alrededor.
Sin embargo, tales animales compiten. Compiten por el espacio
para vivir, ya que tener tal espacio garantiza que tendrán
suficiente comida. Los efectos de la competencia por espacio
son significativos e importantes para los acuaristas de arrecife.
No es competencia por comida...
Los organismos necesitan muchos materiales
diferentes para crecer y reproducirse. Cuando un material
o factor requerido es absolutamente necesario para el crecimiento
o capacidad reproductiva del organismo y ese material es escazo,
entonces se dice que ese material es "limitante."
Sólo cuando un material es limitante simultáneamente
para dos organismos ocurrirá la competencia. Esto debería
ser obvio, ya que si hay más que suficiente de un recurso
alrededor, nadie va a pelear por él. Sólo cuando
la sobrevivencia de uno es amenazada vale la pena pelear por
materiales. Esto se debe a que pelear por materiales tiene
sus costos y los organismos están en una lucha desesperada
por minimizar los costos.
Los no Biólogos a menudo piensan
que una buena definición de evolución es "la
sobrevivencia del más apto," con énfasis
en "sobrevivencia". Todo esto se remonta al viejo
dicho acuñado por un Biólogo victoriano enamorado
con el concepto de evolución de que, "La naturaleza
se tiñe de rojo con la sangre en los dientes y las
garras..." Y siguiendo esa tendencia de pensamiento,
el animal más adaptado es el "sobreviviente."
Pero hay más en adaptación que la simple sobrevivencia.
La adaptación evolucionaria o "Darwiniana"
no es simplemente sobrevivencia; más bien es la producción
de descendencia sobreviviente. Dentro de este contexto, el
organismo "más adaptado" es aquel que produce
la mayor descendencia que sobreviva y se reproduzca en la
siguiente generación. Es por eso que el manejo de los
"costos" es tan importante. Mientras que los acuaristas
de arrecife están familiarizados con el concepto de
presupuesto, pocas veces se dan cuenta que los animales también
tienen un presupuesto.
El presupuesto de nuestros animales no
es en efectivo, sino en energía y materiales. Durante
milenios de evolución, la fisiología de cualqueir
organismo ha sido ajustada a usar la energía y materiales
disponibles de la manera más eficiente. Toda esta energía
y materiales básicos que el animal obtiene, los obtiene
como comida de una u otra forma del ambiente que lo rodea.
Por lo que respecta al animal, la energía en un sentido
básico es azúcar y los materiales básicos
para la construcción de tejidos, proteínas.
La fuente primaria de ambos, energía y material, es
la comida.
Para los animales que se alimentan por
suspensión, en la mayoría de los ecosistemas
marinos de aguas someras , hay suficiente comida disponible
todo el tiempo para cumplir las necesidades básicas
del animal. Las necesidades básicas, en orden, son:
1. Mantenimeinto de tejidos simple o
respiración básica; este es el proceso simple
de seguir con vida. Si el organismos tiene energía
y materiales más que suficientes para seguir con
vida, la siguiente prioridad es:
2. Reparar los daños. Si el organismos tiene energía
y materiales más que suficientes para reparar los
daños, la siguiente prioridad es:
3. Crecer. Si el organismos tiene energía y materiales
más que suficientes para crecer, la siguiente prioridad
es:
4. Reproducción. Si el organismos tiene energía
y materiales más que suficientes para seguir con
vida, reparar los daños y crecer, entonces virtualmente
todo el exceso de energía es puesto en la reproducción.
De hecho, una vez que el tamaño
básico es alcanzado de tal forma que recursos suficientes
puedan colectarse y destinarse a reproducción, la mayoría
de los animales detienen el crecimiento y ponen todo el exceso
de energía en la reproducción. Los tamaños
adultos de la mayoría de los invertebrados marinos
parecen estar determinados por los ajustes necesarios para
obtener suficiente comida para reproducirse.
En el acuario, a menudo la comida puede
considerarse como limitante del crecimiento de los organismos.
A menudo los aficionados alimentan en forma insuficiente o
pueden tener sólo comidas inadecuadas disponibles.
En el mundo real, en contraste con el acuario, la comida nunca
escacea; en el arrecife de coral, probablemente ningún
coral, coral blando o cualquier animal que se alimente por
suspensión estará limitado por la comida. Si
todas las demás cosas están iguales, los animales
tendrán suficiente comida para alcanzar el tamaño
adulto y persistir para reproducirse sin problemas.
Bueno, todas las otras cosas nunca están
iguales. "La mosca en la sopa" no es la falta de
comida sino la falta de espacio.
Para los animales que se alimentan en la
columna de agua en virtualmente todos los ambientes marinos
bénticos, mientras que la comida no es limitante, el
ambiente no es infinito. El factor limitante es, simplemente,
el espacio que ocupan mientras se alimentan y viven.
Como consecuencia, entonces, los animales inmóviles
del fondo que se alimentan por suspensión como corales,
corales blandos, briozoos, tunicados, esponjas, anémonas
y algunos gusanos tubícolas, todos compiten por espacio
(Benayahu and Loya. 1981; Wellington, 1982; Miller, 1998;
Muko, et al, 2001a,b). Es interesante que, estos son los organismos
que el acuarista intenta mantener en su sistema.
Es competencia por espacio…
Los no Biólogos, tales como los
acuaristas de arrecife, a menudo ven la competencia con una
analogía a los deportes o los negocios y a menudo parecen
tener le punto de vista de que una competencia "buena,
saludable" es algo en lo que sus animales deberían
participar y beneficiarse de ello. Bueno, es una bonita idea;
total, absolutamente y 100% errónea, pero aún
una bonita idea. Sería mejor darle un vistazo a la
industria de agricultura. Si quieres cosechas saludables,
las proteges -con un costo razonable- de toda competencia.
La mejor competencia es la NO competencia. En el mundo natural,
la competencia te mata igual que la predación.
En un nivel más básico, la
competencia desvía recursos que pudieran usarse para
otros aspectos de la fisiología de los organismos.
Estos recursos incluyen ambos, energía y materiales.
La competencia alenta el crecimiento, causa daño y
alenta la reparación del daño, y puede parar
toda reproducción. El impacto relativo de de la competencia
es dependiente del tipo de competencia que está ocurriendo,
con el impacto menor proveniente de competencia como resultado
de la forma de crecimiento. Un impacto significativamente
mayor resulta de algunos tipos de competencia más agresivos.
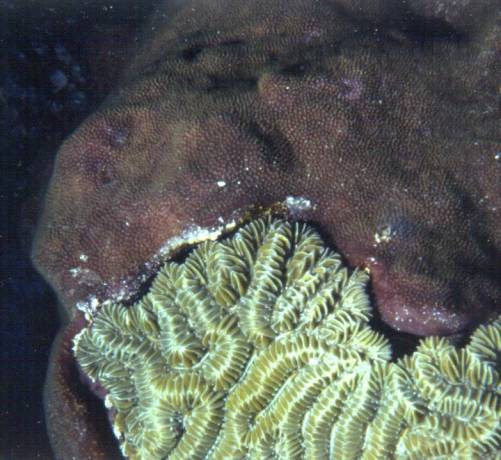
Figure
1. Estas dos cabezas de coral en un arrecife caribeño
están peleando hasta el final, y parece que el de abajo
(Meandrina meandrites), está ganando.
Competencia pasiva vista en el acuario
Probablemente el tipo más básico
de competencia es la competencia pasiva vista cuando un organismo
crece de tal manera para interceptar un recurso limitante.
Para los organismos zooxanthelados como los corales, la luz,
así como el espacio, pueden ser un recurso limitante.
Muchos tipos de corales tienen una forma de crecimiento corporal
básica que les permite excluir otros organismos zooxanthelados
cercanos. Lo hacen sobrecreciendo y sombreándolos.
Este tipo de competencia es todavía competencia por
espacio, ya que el resultado neto es que el ganador es capaz
de mantener su territorio, pero es considerada pasiva. Aún
cuando el ambiente tiene algún efecto en la forma final
del animal, la mayoría de los factores determinantes
son simplemente la genética del organismo y éstos
van a ocurrir si hay o no un competidor potencial presente.
Las formas de crecimeinto de este tipo incluyen los corales
tabulados tales como varias especies de Acropora. Este
tipo de competencia puede ser visto en pequeña escala
en el acuario. Dado que la mayoría de los acuaristas
tienden a mantener su acuario en un presupuesto de deprivación,
la contribución de los azúcares producidos por
las zooxanthelas al presupuesto total de energía del
animal puede ser muy alto. Al mismo tiempo, la intensidad
de la luz en el sistema puede ser realtivamente baja. El resultado
neto de tales condiciones es que algunos organismos zooxanthelados,
quiza algunos corales, pueden estar al borde de la malnutrición.
Tener un organismo creciendo sobre y sombreando un coral y
subsecuentemente causarle su decline o pérdida, es
algo que muchos acuaristas han atestiguado en sus sistemas.
Por supuesto, tal competencia puede ser disminuída
de varias maneras. La forma primaria de prevenir la ocurrencia
de dicha competencia es colocar los corales separados. Trabajo
realizado en la gran barrera de arrecife (Endean, et al. 1997)
indica que aún las cabezas de coral pequeñas
difícilmente están cercanas a otras a menos
de 30 cms. Ese debería ser un buen espacio también
en el acuario.
 |
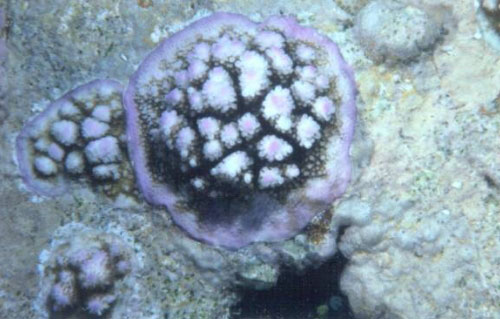
Figure
2. Esta Acropora tabular está "sombreando"
cualquier competidor potencial. Sin embargo, tales formas
de crecimiento son hidrodinámicamente inestables y
podrían romperse durante tormentas o aún con
corrientes mareales fuertes.
Competencia activa, cómo se ve en el acuario
La agresión activa es una proposición
mucho más seria que la pasiva ya que involucra destinar
y gastar recursos que son escazos. La detección de
un competidor potencial cercano es probablemente el estímulo
más importante que cualquier animal sésil puede
procesar. Generalmente, si un depredador es detectado, un
animal sésil tiene pocas opciones. Será atacado
o no, pero independientemente del resultado, él no
puede huir. Algunos animales sésiles pueden producir
estructuras para protegerse o químicos si lo necesitan
y algunos tienen comportamientos defensivos, pero generalmente
los animales sésiles están a merced del depredador
que los descubre (Miles, 1991).
Este no es el caso con respecto a los competidores.
La competencia por espacio generalmente no es un proceso rápido;
ocurre cuando dos animales comienzan a crecer juntos y a menudo
es un proceso relativamente lento. Esto significa que los
competidores pueden preparar sus recursos para una lucha larga
o decisiva. Las estrategias para la pelea son diferentes,
estas diferencias se reflejan en la distribución de
los recursos.
Algunos animales son maestros del ataque
a largo plazo. Generalmente, estos tienden a ser animales
de crecimiento lento; en el mundo de los corales, estos pueden
ser animales que forman colonias masivas. Resultados experimentales
en el Caribe indican que los corales masicvos de crecimiento
lento pertenecientes a las familias Mussidae, Meandrinidae
y Faviidae son las especies más agresivas (Lang, 1973;
Lang, and Chornesky. 1990). Tales animales atacan a sus competidores
usando agentes destructivos tales como cordones mesenteriales,
tentáculos y químicos potentes. Sus recursos
son dirigidos a un forcejeo largo. Intentan matar y sobrecrecer
a sus competidores adyacentes y pueden reponerse del daño
remarcablemente bien. La cara opuesta de esta estrategia es
que ellos crecen muy lento. Muchos de sus recursos parecen
destinados a ser competidores agresivos. Probablemente como
resultado de este tipo de estrategia, las colonias de coral
masivas son animales de largas vidas; se ha demostrado que
algunos tienen miles de años. Es importante reconocer
que la selección natural ha moldeado estos animales
para MATAR a sus competidores potenciales, no para simplemente
desanimarlos. Y, mientras más eficientes sean para
la matanza, más larga y exitosa será su vida.
 |
Figure
3. Estos corales peleaban por espacio en mi tanque hace
algunos años. El Acropora de arriba fue muerto
eventualmente. Noten los cordones mesenteriales extendiéndose
desde la colonia inferior.
Alelopatía: la amenaza impredecible
Se solía pensar que la mayoría
de los corales duros no competían principalmente mediante
químicos, pero a través de los años se
ha acumulado evidencia de que ésta es una suposición
falsa (Gunthorpe and Cameron 1990; vean Borneman, 2002, para
una buena discusión de este tema). El reconocer que
"los llamados" esclaractinios no son un solo grupo
evolutivo definido, sino en realidad una agrupación
superficial resultante de varios linajes evolutivos distintos
y no particularmente relacionados de manera cercana (Romano
and Palumbi, 1996) ayuda a explicar algunas de estas ideas
erróneas. Estudios dentro de cualquier subgrupo de
escleractinios, probablemente no tendrán utilidad para
todo el grupo, ya que el grupo es falso y se basa en criterios
superficiales. Consecuentemente, estudios mostrando una ausencia
de competencia química podrían ser válidos
para una o dos especies, pero no para el grupo en su conjunto.
Es probable que muchas de estas colonias de larga vida y crecimiento
lento estén utilizando muchos de sus recursos para
producir químicos que son asesinos potentes de competidores
potenciales. En la naturaleza, la producción de éstos
podría inhibir naturalmente el asentamiento y la metamorfosis
de larvas de coral cerca de la colonia, o podrían simplemente
matar colonias pequeñas de corales creciendo cerca
de una colonia grande. Tal guerra química es llamada
alelopatía, y es bien conocida entre los corales blandos,
esponjas y tunicados. Es probablemente una estrategia importante
también entre algunos de los corales duros (Borneman,
2002).
En un acuario, la producción total
de químicos alelopáticos tóxicos podría
ser bastante pequeña. Sin embargo, tales materiales
tienden a ser excepcionalmente tóxicos y en los confines
de un acuario pequeño, digamos cualquier cosa más
pequeña que un par de miles de galones, la presencia
o ausencia de corales produciendo tales químicos podría
realmente determinar el éxito del sistema completo.
La magnitud de los efectos de los químicos alelopáticos
es absolutamente impredecible dado que no tenemos ninguna
manera de probarlos o medirlos porque:
1. Tenemos poca información básica
acerca de cuáles corales duros los utilizan.
2. Casi no tenemos información acerca de qué
corales son afectados por ellos.
3. No tenemos información acerca de qué dispara
la producción de los químicos.
El último factor mencionado arriba
puede ser el más crítico. Colocar un fragmento
de coral nuevo cerca de un fragmento ya establecido podría
ser el análogo de establecimiento y crecimiento de
un coral juvenil situado cerca y podría disparar la
producción de los químicos y esto podría
tener resultados desastrosos para todo el sistema.
Figure
4. Noten la colonia central sobrecreciendo la colonia
pequeña de la izq. Las colonias más grandes
tienen más recursos para dedicarle a la competencia
y a menudo ganan en sus peleas. La ausencia de colonias en
esta área, aproximádamente a 30 cms a un lado,
es debida probablemente a la presencia de químicos
alelopáticos de cabezas de colonias más grandes
cercanas. Estos químicos inhiben la introducción
de la mayoría de las larvas en esa área.
Ataque físico
Otros corales compiten sin químicos
o utilizan medios adicionales para matar a sus competidores
potenciales. En el acuario, probablemente los más importantes
de estos medios son el desarrollo de estructuras especializadas,
llamadas tentáculos "barredores", usados
de forma agresiva por muchos corales como Galaxea y
Euphyllia. Alguna anémonas también utilizan
tentáculos similares (Richardson, et al., 1979; Williams,
R. B. 1991; Hidaka et al. 1997; Langmead y Chadwick-Furman.
1999a,b). Hace aproximadamente unos 20 años, el papel
de estos tentáculos como estructuras agresivas no había
sido descubierto y se suponía que eran un medio para
obtener comida extra. Si se toman un tiempo para leer algo
de la literatura científica antigua de anémonas
y corales, se podrían encontrar con el término
de "tentáculo de captura" empleado para describir
estas estructuras, ya que se pensaba servían en alguna
función especial para la "captura" de comida.
Bueno...ellos no capturan comida, se usan para matar incrustadores
ofensivos en el espacio del coral o la anémona y pueden
hacer un muy buen trabajo. Estos tentáculos especializados
contienen un armamento más potente de nematocistos,
en número y tipo, que los tentáculos regulares.
Lo que considero interesante de tales tentáculos es
su absoluto y mayor tamaño. En muchos Euphyllia
estos tentáculos pueden alcanzar de 30 a 40 cms de
largo y son más o menos transitorios. El animal los
extiende a voluntad, y esto podría ser cuando el aficionado
no está viendo su tanque. La primera vez que estas
estructuras se ven, puede ser ilustrativo y atemorizante.
 |
Figure
5a. Un pequeño parche clonal de la anémona
temperada Metridium senile (los individuos grises,
centro-arriba) dentro de un ensamble clonal de Metridium
giganteum blanca y mucho más grande, fotografiado
en Puget Sound. Los individuos de ambas especies se
alimentan por suspensión. La única forma
en que Metridium senile puede persistir en esta
área es agrediendo actívamente a su primo
más grande.
|
 |
Figure
5b. Uno de los individuos de Metridium senile,
mostrando sus tentáculos agresores. Noten cuan
grandes son estos tentáculos blancos, cargados
de nematocistos, comparados con los finos tentáculos
filamentosos para alimentación.
|
Mi primera experiencia con los tentáculos
barredores vino poco después de que tuve mi primer
Euphyllia, hace 15 años. Estaba familiarizado
con tales estructuras, en teoría; había leído
acerca de ellos, pero nunca los había visto "en
acción." Traje este bello coral a casa de la tienda,
y lo instalé en mi tanque. Todo iba bien por un rato.
Un par de días después, llegué a casa
del trabajo, agarré una lata fría de cerveza
del refri y me senté enfrente de mi tanque. Y entonces
observé como esta estructura larga, delgada, se elevó
de mi coral y bastante deliberadamente comenzó a moverse
alrededor del coral. Fue realmente fascinante. Aún
más facinante fue la respuesta de los animales que
tocó. El tentáculo tocó delicadamente
el manto de una Tridacna que estaba cerca del coral
y en cada punto de contacto apareció una mancha blanca.
En el lapso de una hora, cada una de esas manchas era un hoyo
en el manto. Donde los tentáculos tocaron unos zooántidos,
estos pólipos simplemente se contrajeron y nunca volvieron
a abrirse. Donde tocaron otro coral, bueno, eso fue todavía
más interesante, porque parecía provocar una
reacción de hinchamiento, pero no parecía matarlo,
no al menos al primer contacto. No tuvo una segunda oportunidad.
Después de acabarme mi líquido espumoso (uno
debe tener las prioridades adecuadas), reacomodé mi
tanque para que mi coral no tuviera la oportunidad de "estirarse
y tocar a alguien."
La agresión competitiva en el acuario,
por lo tanto, puede tener 3 manifestaciones; una es específica,
de rango relativamente corto y determinada por qué
tan lejos puede el animal alcanzar a hacer daño. La
segunda es un sistema más amplio y los efectos son
determinados por la potencia de los químicos alelopáticos
y su abundancia. El método final podría ser
un proceso mucho más lento tal como el sobrecrecimiento
físico.
Las respuestas a estos ataques pueden variar,
pero al menos, la detoxificación de los químicos
consume energía del receptor y ésto es suponiendo
que los químicos pueden detoxificarse. La reparación
de los daños causados por los tentáculos barredores
y otras estructuras agresivas tales como los filamentos mesenteriales,
es más directa, pero requiere materiales estructurales
tales como las proteínas y energía de la comida.
Prevención de interacciones competitivas
en el acuario
Juzgando por la revisión de varios
"tanques del mes" creo que es probablemente infructuoso
sugerir que la mayoría de los aficionados traten de
evitar la competencia en sus sistemas. Muchos de estos sistemas
son "tanques de poster" para la competencia en acción
y este tipo de tanque es visto contínuamente como "la
manera de hacerlo." Sin embargo, muchos de estos tanques
contienen versiones Bonsai de corales colocados "cachete
con papada" (si los corales tuvieran mejillas o papadas)
básicamente le parecen a este científico como
un experimento de competencia corriendo desordenadamente.
La gracia que salva estos tanques es que algunos de los corales
en ellos, tales como muchos de los Acropora, a menudo
no parecen ser competidores particularmente buenos o efectivos
usando químicos o defensas urticantes. En la naturaleza,
muchos parecen llenar el nicho ecológico de las "hierbas;"
esto es, llegan a un sistema, crecen rápido y se reproducen
y entonces son reemplazados por animales más exitosos,
pero de crecimeinto más lento que los derrotan en la
competencia por espacio. Básicamente estos corales
están adaptados a vivir en situaciones donde el ambiente
está en estado continuo de disrupción, que en
la naturaleza es provocado principalmente por las tormentas.
Estos corales prosperan en la disrupción continua de
los acuarios de arrecife, y dado que no son competidores particularmente
efectivos, persisten donde competidores de vidas más
largas y mejores podrían estar sufriendo severamente
de claustrofobia competitiva.
Muchos aficionados parecen escoger estos
corales potencialmente malos competidores para sus sistemas.
Estas elecciones juiciosas, o más bien accidentales
de animales que son probablemente competidores relativamente
malos son una forma simple, e inconsciente de mantener las
presiones competitivas en el acuario de arrecife en un nivel
bajo. Muchos de los mejores competidores, tales como algunas
especies de los Euphyllia, tienen la reputación
de ser difíciles de mantener, probablemente por su
naturaleza competitiva. Adicionalmente, como a casi todos
los aficionados les agrada ver crecimiento rápido,
tienden a escoger corales que crecen rápidamente. Los
de crecimiento lento y coincidentemente, productores prolíficos
de químicos alelopáticos simplemente no son
elegidos por muchos aficionados. Sin embargo, si se les escoge,
tales corales pudieran, con el tiempo, encontrar situaciones
en el ambiente altamente "empacado" del tanque del
aficionado que estimule la producción de venenos en
cantidades que permitan su acumulación en nuestros
sistemas. En tales situaciones, estos corales probablemente
acabarán envenenándose a sí mismos así
como al resto del sistema. La relativa facilidad con la que
muchos de los corales de bocas pequeñas pueden ser
mantenidos, podría simplemente ser un reflejo de su
naturaleza "herbácea", no competitiva, que
los acuaristas han adaptado para su beneficio.
|