|
Introduction:
As a professor teaching invertebrate zoology,
one of the questions I tell my students that they need to
answer about every animal they examine is, "How does
it feed?" Feeding is the primary motivator for animals.
Even though some animals will go without food for a time for
reproductive related behaviors, if the animal cannot feed,
everything else is immaterial; without food it will die. Differences
in the morphology of the feeding apparatus are some of the
basic ways in which animals differ, and are reflected in the
many and varied body forms that animals take.
People are accustomed to thinking of animals
as mobile creatures, moving from place to place. Generally,
when they think about these movements, most people will realize
the movements are due to the animals' requirements for food.
Herbivores move around as they exhaust the tasty plants in
any one place, and predators move following their moving prey.
But, what about animals that don't have to chase down food?
How does the "need to feed" get manifested in changes
in their behavior or morphology? Often the changes show up
in other aspects of their natural history. Many good examples
of that are found in the suspension-feeding animals of shallow
water marine environments such as coral reefs.
For animals living in the shallow water
marine environments, food is almost never in short supply
(a fact that is very important to reef animal husbandry and
which has been commented on numerous times in the pages of
this magazine - see previous articles (1,
2,
3,
4,
5,
6,
7)
by Eric Borneman and myself.
The amount of life, of all shapes and sizes, in the planktonic
environment is well beyond easy comprehension. Additionally,
shallow oceanic waters are always in motion, so this life
is also continually being moved around, and it may also be
moving of its own accord.
This means that the water in shallow oceanic
environments, down to a depth of a few hundred feet or so,
is full of small living organisms. Small living organisms
are always food for some other organisms, so basically the
water in this layer of the oceans is a mobile, continually
self-renewed snack bar waiting to be harvested. For an animal
that can utilize some method to glean food from the water,
the food is never in short supply; each wave, current or eddy
brings a new fresh supply of goodies to be eaten.
Not surprisingly, then, natural selection
has favored the development of a lot of ways to harvest this
food. There are various types of passive or active suspension
feeding animals as well as animals that might be considered
to be more actively predatory on specific components of the
water borne array of foods. Such animals are generally not
too selective about their food. As long as it floats or swims,
they will try to eat it. Probably because of this general
non-selectivity, hobbyists often find that these animals are
relatively easy to keep alive, at least for a while. If some
relatively nutritious food is placed in the water the animals
will feed on it.
Consequently, most animals that reef aquarium
hobbyists keep fall into the category of animals that feed
directly or indirectly off the plankton. What these suspension-feeding
animals don't do, however, is compete for food. Normally,
for these kinds of animals, there is always enough food to
go around. However, such animals do compete. They compete
for the space to live, because having that space guarantees
that they will get enough food. The effects of that competition
for space are both significant and important for reef aquarists.
Competition For Food It Isn't…
Organisms need a lot of different materials
for growth and reproduction. When an essential material or
required factor is absolutely necessary for the growth or
reproductive capacity of organisms, and that material is in
short supply, then that particular material is said to be
"limiting." Only when a resource is simultaneously
limiting for two or more organisms will competition occur.
This should be obvious, for if there is more than enough of
the resource to go around, nobody will fight over it. Only
when one's survival is at stake is it worth it to fight for
materials. This is because fighting for materials has its
costs, and organisms are in a desperate struggle to minimize
costs.
Non-biologists often think that a good
definition of evolution is "Survival Of The Fittest,"
with the emphasis on "Survival." This all harkens
back to the old saying, coined by a Victorian biologist enamored
with the concept of evolution that, "Nature was red with
blood in tooth and claw…" And following this train
of thought, the fittest animal is the "survivor."
Well, to be sure an animal has to survive to be "fit."
But there is more to fitness than simple survival.
Evolutionary or "Darwinian" fitness
is not simple survival; rather it is the production
of surviving offspring. Within any given context, the "fittest"
organism is the one that produces the most offspring that
survive to live and reproduce in the next generation. This
is why management of "costs" is so important. While
reef aquarists are often very familiar with the concept of
a budget, they seldom seem aware that their animals are also
on a budget.
The budget that our animals are on is not
one of cash, but one of energy and materials. Over millennia
of evolution, any organisms' physiology has been fine-tuned
to utilize the energy and material that is available to it
in a most efficient manner. All of this energy and all of
the raw materials that an animal gets, it gets as food in
one form or another from the environment surrounding it. As
far as animals are concerned, energy in the basic sense is
sugar, and the raw material for tissue construction is protein.
The primary source for both energy and material is feeding.
For suspension-feeding animals, in most
shallow water marine ecosystems, there is sufficient food
available at all times to fulfill, the animals' basic needs.
These basic needs, in order, are:
1. Simple tissue maintenance or basic
respiration; this is the simple process of staying alive.
If the organism has more than enough energy and materials
to stay alive, the next priority is:
2. Repair of injuries. If the organism has more than enough
energy and materials to stay alive and repair injuries,
the next priority is:
3. Growth. If the organism has more than enough energy and
materials to stay alive, repair injuries, and to grow, the
next priority is
4. Reproduction. If the organism has enough energy and materials
to stay alive, repair injuries and to grow, then virtually
all the excess energy is put into reproduction.
In fact, once a basic size is reached so
that sufficient resources can get collected and be allocated
for reproduction, most animals cease much growth and put all
of their excess energy into reproduction. The adult sizes
of most marine invertebrates appear to be determined by the
tradeoffs necessary to obtain sufficient food to reproduce.
In aquaria, food may often be considered
to limit the growth of organisms. Often, hobbyists feed insufficient
amounts of food or may only have inappropriate foods available
to them. In the real world, as opposed to reef aquaria, food
is never in short supply; on a coral reef, no coral, soft
coral, or any suspension-feeding animal is likely to be limited
by food. All other things being equal, the animals will have
enough food to reach an adult size and persist to reproduce
without any problems.
Well…. All other things are never
equal. The "fly in the ointment" is not lack of
food, but lack of space.
For animals feeding from the water column
in virtually all marine benthic environments, while food is
not limiting, the environment is not infinite. The limiting
factor is, simply, the space to occupy whilst you feed,
and live. It follows, then, that immobile bottom-dwelling,
suspension-feeding, organisms such as corals, soft corals,
bryozoans, tunicates, sponges, sea anemones, and some tube
worms all compete for space (Benayahu and Loya. 1981; Wellington,
1982; Miller, 1998; Muko, et al, 2001a,b). Interestingly enough,
these are just the sorts of animals reef aquarists like to
maintain in their systems.
Competition for Space It Is…
Non-biologists, such as reef hobbyists,
often view competition with a sports or business analogy,
and often seem to have the viewpoint that "good, healthy
competition" is something that their animals should be
able to engage in and thrive from it. Well, this is a neat
idea; totally, absolutely and 100 percent wrong, but still
a neat idea. It would be better to take a look at the agriculture
industry. If you want good healthy crops, you protect them
- at all reasonable cost - from all competition. The best
competition is NO competition. In the natural world, competition
kills you just as dead as does predation.
On the most basic level, competition diverts
resources that might be used for other aspects of the organism's
physiology. These resources include both energy and materials.
Competition slows growth, causes injury and slows injury repair,
and may stop reproduction altogether. The relative impact
of competition is dependent on the type of competition occurring,
with the least impact occurring from competition resulting
from simple growth forms. Significantly greater impact occurs
from competition resulting from more aggressive types of competition.
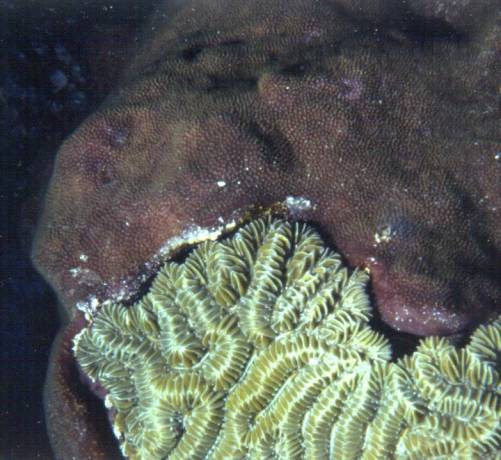
Figure
1. These two coral heads on a Caribbean reef are fighting
to the finish,
and it looks like the lower one (Meandrina meandrites)
is winning.
Passive Competition Seen In Aquaria
Probably the most basic type of competition
is the passive competition seen when one organism grows in
such a manner as to intercept a limiting resource. For zooxanthellate
organisms such as corals, light, as well as space, may be
a limiting resource. Many types of corals have a basic body
growth form that allows them to exclude other nearby zooxanthellate
organisms. They do this by overgrowing and shading them out.
This type of competition is still competition for space, as
the net result is that the winner is able to hold on to its
turf, but it's considered to be relatively passive. Even though
the environment has some effect on the animal's final shape,
most of the determining factors are simply the genetics of
the organism and these both would occur whether or not a potential
competitor was present. Growth forms such as this include
tabulate corals such as several species of Acropora.
This type of competition may be seen on a small scale in many
aquaria. Because most aquarists tend to keep their aquaria
on a starvation budget, the contribution of the sugars produced
by zooxanthellae to the overall energy budget of the animal
may be quite high. At the same time, light intensity in the
system may be relatively low. The net result of such conditions
is that some zooxanthellate organisms, perhaps some corals,
may be on the thin edge of malnutrition. Having one organism,
growing over and shading such a coral and subsequently causing
its decline or demise, is something that many aquarists have
witnessed in their systems. Such competition can be ameliorated
in any number of ways, of course. The primary way to prevent
such competition from occurring would be to initially place
the corals further apart. Work done on the Great Barrier Reef
(Endean, et al. 1997) indicates that even small coral heads
are seldom closer to one another than about one foot. That
would be good spacing in an aquarium as well.
 |
Figure
2. This tabulate Acropora is "shading out"
any potential competitors. However, such growth
forms are hydrodynamically unstable, and may break up during
storms or even high tidal currents.
Active Competition, As Seen In Aquaria
Active aggression is a much more serious
proposition than is passive aggression as it involves the
allocation and expenditure of scarce resources. The detection
of a nearby potential competitor is probably the most important
sensory input that any sessile animal can process. Generally,
if a predator is detected, a sessile animal has few options.
It will either get attacked or it won't, but whatever the
outcome, it can't flee. Some sessile organisms can produce
protective chemicals or structures on demand, and some have
defensive behaviors, but generally sessile animals are simply
at the mercy of a predator that discovers them (Miles, 1991).
This is not the case with regard to competitors.
Competition for space is generally not a rapid process; it
occurs as two animals tend to grow together, and this is often
a relatively slow process. This means that the competitors
can take the time to marshal their resources for a long or
decisive fight. The strategies for fighting often differ,
and this difference is reflected in the allocation of resources.
Some animals are masters of the long-term
attack. Generally, these tend to be slow growing animals;
in the coral world, these would be animals that form massive
colonies. Experimental results in the Caribbean indicate that
the slow-growing massive corals belonging to the families
Mussidae, Meandrinidae, and Faviidae are the most aggressive
species (Lang, 1973; Lang, and Chornesky. 1990). Such animals
attack their competitors using destructive agents such as
mesenterial strands, tentacles and potent chemicals. Their
resources seem allocated for the long-haul. They actively
attempt to kill and overgrow adjacent competitors, and they
can heal injury remarkably well. The flip side of this strategy
is that they grow very slowly. Many of their resources appear
to be allocated to being aggressive competitors. Probably
as a result of this sort of strategy, massive coral colonies
are some of the longest lived of animals; some have been shown
to be several thousand years old. It is important to realize
that natural selection has fashioned these animals not to
simply "discourage" any potential competitor, but
instead to KILL the potential competitor. And, the more efficient
they are at killing it, the longer and more successful their
life will be.
 |
Figure
3. These corals were fighting for space in one of my aquaria
several years ago. The Acropora,
at the top, was eventually killed. Notice the mesenterial
strand extending from the lower colony.
Allelopathy: The Unpredictable Menace
It used to be thought that most stony corals
did not primarily compete with chemical means, but evidence
has been accumulating for several years that this is a false
assumption (Gunthorpe and Cameron 1990; See Borneman, 2002,
for a good discussion of this topic). The realization that
"so-called" scleractinian corals are not a single
evolutionarily defined group, but actually a superficial grouping
resulting from several distinct and not-particularly closely
related evolutionary lineages (Romano and Palumbi, 1996) helps
explain some of the misconceptions. Studies within any subgroup
of the scleractinians, probably while not have utility throughout
the group, as the group is a false one based on superficial
criteria. Consequently, studies showing a lack of chemical
competition might be valid for one or a few species, but would
not be valid for the group as whole. It is likely that many
of these slowly growing, and very long-lived colonies are
utilizing a lot of their resources to produce chemicals that
are quite potent killers of potential competitors. In nature,
the production of these chemical factors would naturally inhibit
the settlement and metamorphosis of coral larvae near to the
colony, or they might simply kill small coral colonies growing
near the larger colony. Such chemical warfare is called allelopathy,
and is well known amongst the soft corals, sponges, and tunicates.
It is likely an important strategy amongst some of the stony
corals as well (Borneman, 2002).
In an aquarium, the absolute production
of toxic allelopathic chemicals would likely be quite small.
However, such materials tend to be exceptionally toxic and
in the confines of a small aquarium, say anything smaller
than a couple of thousand gallons, the presence or absence
of corals producing such chemicals could really determine
the success of the entire system. The magnitude of the effects
of allelopathic chemicals is absolutely unpredictable as we
have no way to test for or measure them because:
1. We have little basic information
about which stony corals use them.
2. We have almost no information about which corals are
affected by them.
3. We have no information about what triggers the production
of the chemicals, and
The last factor above may be the most critical.
Placement of a new coral fragment next to a more established
fragment would be the analogue of settlement and growth of
a juvenile coral settling nearby, and would trigger the production
of chemicals and this could have disastrous results for the
whole system.
Figure
4. Notice the central colony overgrowing the smaller colony
to the left. Larger colonies have
more resources to devote to competition and often win their
competitive encounters. The lack of
other colonies in this area, about a foot on a side, is probably
due to the presence of allelopathic
chemicals from nearby larger coral heads. These chemicals
inhibit recruitment into the
area from most larvae.
Physical Attack
Other corals compete without chemicals,
or utilize additional means of killing their potential competitors.
In aquaria, probably the most important of these means are
specially developed structures, called "sweeper"
tentacles, used in an aggressive manner by many corals such
as Galaxea and Euphyllia. Some anemones also
utilize similar tentacles (Richardson, et al., 1979; Williams,
R. B. 1991; Hidaka et al. 1997; Langmead and Chadwick-Furman.
1999a,b). About twenty or so years ago, the role of these
tentacles as aggressive structures had not been elucidated,
and they were often presumed to be functional as a means of
getting extra food. If you take the time to do some reading
in the older sea anemone and coral scientific literature,
you may come across the term of "catch tentacle"
used to describe these structures as they were thought to
serve some special function in "catching" food.
Well… they don't catch food, they are used to kill offending
encroachers on the space of the coral or anemone, and they
can do a pretty good job of it. These specialized tentacles
contain a more potent armament of nematocysts, both in types
and numbers than do regular tentacles. What I think is interesting
about such tentacles is their absolute, and large, size. In
many Euphyllia these tentacles may reach 12 to 15 inches
in length, and they are more-or-less transitory. The animal
will extend them out at will, and this may not be when the
hobbyist is watching their tanks. The first time such structures
are seen, it may be both enlightening and frightening.
 |
Figure
5a. A small colonal patch of the temperate anemone,
Metridium senile (the gray individuals, upper
center) within a clonal assemblage of the much larger,
white, Metridium giganteum photographed on a
piling in lower Puget Sound. Individuals of both species
are suspension-feeding animals. The only way Metridium
senile can persist in this area is by actively aggressing
against its much larger cousin.
|
 |
Figure
5b. One of the Metridium senile individuals,
showing its aggressive tentacles. Note how much larger
these white, nematocyst laden tentacles are, compared
to the fine filamentous feeding tentacles.
|
My first experience with sweeper tentacles
came shortly after I got my first Euphyllia, about
15 years ago. I was familiar with such structures, in the
abstract; I had read about them, but had never seen them "in
action." I brought this beautiful coral home from the
store, and set it up in my tank. All was well for a while.
A couple of days later, however, I came home from work, grabbed
a cool can of suds from the fridge and sat down in front of
my tank. I then got to watch as this long, thin, structure
rose out of my coral and quite deliberately started to be
moved around the coral. It was truly fascinating. Even more
fascinating was the response of the animals that it touched.
The tentacle gently touched the mantle of a Tridacna
that was near the coral, and at each point of contact, a white
spot appeared. Within an hour, each of those spots was a hole
through the mantle. Where the tentacle touched some zoanthid
polyps, those polyps simply contracted, and never opened again.
Where it touched another coral, well, that was even more interesting,
because it seemed to elicit swelling and inflation responses,
but it didn't seem to kill it, at least not after the first
contact. It didn't get a second chance. After finishing my
can of foamy liquid (one must have the proper priorities),
I rearranged my tank so that my coral didn't have the opportunity
to "reach out and touch someone."
Competitive aggression in aquaria, then,
may have three main manifestations; one is specific, relatively
short-ranged, and determined by how far the animal can reach
to do its damage. The second is more system wide, and the
effects are determined by the potency of the allelopathic
chemicals and their abundance. The final method would be by
much slower processes such as physical overgrowth.
The responses to these attacks can vary,
but at the very least, the detoxification of the chemicals
takes energy away from the recipient, and this is presuming
that the chemicals can be detoxified. Repair of the injuries
caused by sweeper tentacles, and other aggressive structures
such as mesenterial filaments, is more straight forward, but
it takes both structural materials such as proteins and food
energy.
Prevention of Competitive Interactions in Aquaria
Judging by looking back through the various
"Tanks of the Month," I feel that it is probably
fruitless to suggest that most hobbyists should try to avoid
competition in their systems. Many of these systems are "Poster
Tanks" for competition in action, and this kind of tank
seems to be continually thought of as "the way to go."
Nonetheless, such tanks containing many Bonsai'd versions
of corals placed cheek-to-jowl (if corals had cheeks or jowls)
basically looks to this scientist like an experiment in competition
run amok. The one saving grace of many of these systems is
that some of the corals in them, such as many of the Acropora,
often do not seem to be particularly good or effective competitors
using chemicals or stinging defenses. In nature, many of them
seem to fill the ecological niches of "weeds;" that
is, they come into a system, grow up fast and reproduce, and
then they get replaced by more successful, but slower growing
animals which outcompete them for space. Basically such corals
are adapted to living in situations where the environment
is in a continual state of disruption, which in nature is
largely accomplished by storms. These corals thrive in the
continual disruption of reef aquaria, and as they are not
particularly accomplished competitors, they persist where
more long-lived and better competitors would be severely suffering
from competitive claustrophobia.
Many hobbyists seem to choose these potentially
poorly competitive corals for their systems. Such judicious
or, more likely, accidental choices of animals that are probably
relatively poor competitors are a simple, and unconscious
means of maintaining the competitive pressures in reef aquaria
at a low level. Many of the better competitors, such as some
of the Euphyllia species, have the reputation of being
difficult to keep, probably because of their competitive natures.
Additionally, as almost all hobbyists like to see rapid growth,
they tend to choose corals that grow rapidly. The slower growing
and, coincidentally, prolific producers of allelopathic chemicals
are simply not chosen by many hobbyists. If they are chosen,
however, such corals may, in time, encounter situations in
the close-packed environment of a hobbyist's tank that stimulate
the production of poisons in quantities that allow their accumulation
in our systems. In such situations, these corals are likely
to end up poisoning themselves as well as the rest of the
system. The relative ease by which many of the smaller mouthed
corals may be kept may simply be a reflection of their weedy,
non-competitive nature, which aquarists have adapted to their
benefit.
|