|
paradigm (PAR-a-dime):
1)
something serving as an example or model of how things should
be done.
2)
$ 0.20…
Food Availability For Reef Animals
It has been known for some time that corals
and other coral reef animals must feed. Eric Borneman has
been discussing the various aspects of feeding in corals for
the last several months in his column in this magazine, and
some of those data are applicable to reefs in general. Nonetheless,
it is hard to get a good handle on the feeding dynamics of
many reef animals. Interestingly enough, however, what we
do know about feeding on a reef, as a whole, indicates that
aquarists, generally, go about feeding their animals in the
wrong way and with the wrong foods. By examining the types
of available food, and the processes of feeding on a reef,
I think it will become apparent that many of the problems
we have with reef aquaria, such as excess nutrients, excessive
growth of undesirable algae, and the inability to keep some
animals alive and healthy is simply due to the feeding of
inappropriate foods, compounded by feeding in the wrong manner.
Before I can discuss the major points of
this proposition, however, I need to "set the stage"
and discuss feeding in general, and explore a bit of what
we know of reef food webs. Some of this information is the
result of basic biological investigations, while other data
come from some very nicely done ecological studies on the
Great Barrier Reef.
First Point: Animals Need To Feed.
All organisms need some sort of food; in
fact, the ability to either feed or generate one's own food
is probably a pretty decent criterion for describing life.
Many organisms are photosynthetic; in other words, they absorb
light and use that light energy to make chemicals. This production
of chemicals using light energy is limited to those organisms
that have chlorophyll in their bodies: photosynthetic bacteria
(cyanobacteria), many types of algae, and plants. Photosynthetically-derived
chemicals, which are mostly sugars, may be broken down, thereby
releasing a portion of that absorbed light energy as chemical
energy.
This secondary production of chemical energy,
either in the initial organism that produced the sugar or
in some other organism, is the basis for all basic energy
utilization in all organisms. It is done by essentially the
same chemical processes in all organisms, and this is one
of the major reasons to consider all life as related. This
process is called "respiration" and unlike the process
that we normally associate with the term respiration, it really
has nothing directly to do with breathing or gas exchange.
Instead, it refers to the chemical breakdown and oxidation
of sugar to release useable chemical energy in a cell.
The fact that this production of useable
energy from the sugar is independent of the production of
the sugar has some rather profound consequences. Because of
this independence, the sugar may be broken down and used some
distance away from where it is made and some time after it
is made. This means that the organism that uses the sugar
doesn't have to be the one that makes it. In other words,
the development of the cellular biochemistry that allowed
for the indirect utilization of light energy, allowed for
the development of animals. Animals are organisms that don't
have the photosynthetic machinery which is characteristic
of the plants, algae, or cyanobacteria that comprise the primary
producer trophic level in ecosystems. They must consume their
food, and because of this they must either eat plants, plant
byproducts, or other animals.
|
|
|
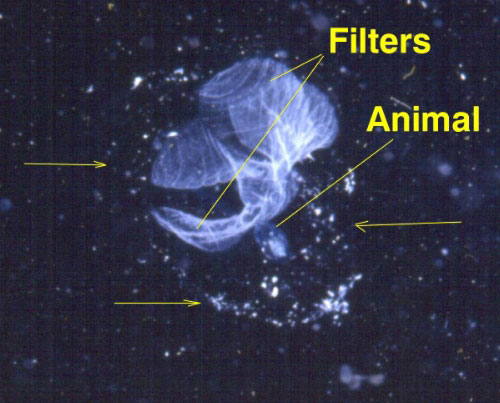
Figure
1. Most reef animals are eating small zooplankton
that are gelatinous in nature. This is a photo of a
larvacean tunicate called Oikopleura. Unlike
benthic tunicates, or sea squirts, larvaceans spend
their entire lives as planktonic animals. They construct
a transparent "house" (the edges of which
are indicated by the arrows) out of mucus secreted by
the epidermis of the animal's head. This house is about
¼ inch in diameter. In the house are the filters
they use to collect food. The animal pumps water through
the filters, and when they are full, the animal eats
them, abandons the house and secretes a new house. It
may do this several times an hour. Larvaceans, and old
larvacean houses, are some of the most common, and nutritious
of the gelatinous zooplankton (See Alldredge, 1972).
|
Second Point: Animals Need To Feed On Organisms;
They Don't Just Live On Photosynthetic Byproducts.
Photosynthesis produces only sugars. It
is the process of using light energy to fuse six carbon dioxide
molecules and six water molecules together to make a simple
sugar. In doing so, it gives off six oxygen molecules as a
waste byproduct. Sugar is called a carbohydrate, because it
contains only carbon, hydrogen and oxygen. Carbohydrates are
useful and necessary chemicals; they may be burned for fuel,
converted to fats or starches to store fuel, or fashioned
into long chains as structural molecules such as cellulose
and chitin. What they can't do, however, is be used to directly
make a protein.
Proteins are the building blocks of all
animal tissues, and the major components of all cells in all
organisms. They are made of subunits called amino acids, often
hundreds of them, fastened together in long chains. Amino
acids, as their name implies, are molecules that have both
an acid and an ammonia residue attached to them. Over 150
amino acids are found in nature, but the vast majority of
proteins are made from only about 20 of them. This large number
of amino acids may be hooked together in an almost endless
variety of ways. And animal chemistry can build, remodel,
and modify proteins wonderfully well. What animal chemistry
cannot do is synthesize an ammonia group from nitrogen and
hydrogen, nor can animals utilize nitrate or nitrite to form
ammonia. This synthesis is largely done by bacteria or photosynthetic
organisms.
Animals cannot manufacture amino acids
from such basic chemical constituents as an ammonia or amine
group and an organic acid, consequently, they must get them
from some other source. Coral reef animals have one or two
options for obtaining their amino acids. If they have zooxanthellae,
they may get some amino acids from the zooxanthellae. Unfortunately,
this is a zero-sum situation. As the zooxanthellae live within
their host, any ammonia that they can utilize must come from
their hosts' tissues as a waste product. If such ammonia is
a waste product of the host, it is largely a byproduct of
the host's metabolism or digestion. This means that the hosts
always will require more amino acids, by a very large margin,
than the zooxanthellae can provide. What the zooxanthella
may do, however, is provide particular types of nitrogenous
products unavailable elsewhere. However, even so, zooxanthellate
animals must be getting their nitrogenous chemicals from another
source, and that source is from feeding of one sort or another.
Animals without zooxanthellae will not, of course, have this
option. They simply must fulfill all of their needs from feeding.
Marine animals typically require that between
five percent and 60 percent of the dry weight of the diet
must be protein. For optimal growth of fish, the diet must
be from 30 percent to 60 percent, depending on the fish. The
absolute requirement from most inactive invertebrates is toward
the lower end of the range, but for highly active invertebrates
such as squids, it is likely as high as fish. All of this
protein must come from either eating some other animal, alga,
or plant; direct absorption from the water around the animal,
or from a zooxanthellate symbiont. Direct absorption of dissolved
amino acids is typically efficiently done in most marine invertebrates,
however, there really is very little of this material available
in natural systems. In a coral reef aquarium, however, this
may be major source of amino acid accumulation by many animals.
Production of amino acids by a zooxanthellate symbiont is
of limited value, as most animals require a far larger amount
of amino acids than may be available from this source. However,
this latter source may provide some essential materials. Most
amino acids, however, probably come from the assimilation
of foods, including bacteria. Bacteria, in fact, are an important
food for most benthic or bottom-dwelling marine animals. This
is because bacteria have higher nitrogen to carbon ratios
in their cells than do either typical animals, plants or algae.
As a consequence many marine animals are specialized to eat
bacteria, either directly out of the water column or indirectly
as a frosting on sediment or detritus particles.
Protein is often a critical resource for
animals. Farmers and aquaculturists have long known that one
way to get maximum growth in captive animals is to make sure
that they have access to a high protein diet. Such diets promote
rapid growth and seem to foster generally good health in animals.
Unfortunately, such diets are quite unnatural in coral reef
areas.
|
|
|
Figure
2. One other common member of the gelatinous zooplankton
food category of coral reef animals is yet another type
of pelagic tunicate called a "salp." Salps
are much like the benthic sea squirts, except that they
live totally in the plankton as mobile colonial animals.
They form long chains of individuals fastened together
at the sides of their tunics. They move through the
water propelled by the water they suck through themselves
to filter it. This is actually a type of jet propulsion.
Salp colonies may be huge. I have seen salp chains,
colonies really, consisting of many thousands of individuals,
in excess of 70 feet long, moving through the water.
In this photo, the guts of the animals are clearly evident
and the edges of the bodies are faintly visible. Each
individual here is about an inch wide. Both living and
dead salp individuals and chains are also common foods
of reef animals, including fishes (see Figure 4).
|
Some Data From Real Reefs
Good data about feeding in corals is relatively
sparse and surprisingly diffuse. There are a few studies from
the Caribbean, a few from the Red Sea, a few from here and
a few from there. There is no good coherent body of knowledge
regarding the types of foods corals and coral reef animals
actually eat, and in what quantities. And, we really have
few data on what foods and in what quantities are necessary
to keep corals and coral reef animals in good health. There
is a real good reason for this lack of information. Although
it is conceptually easy to visualize how to collect such feeding
data, in practice collecting such data becomes very difficult.
The food is often small and fragile; the animals tend to regurgitate
when handled. Additionally, these animals often digest the
food very swiftly. This means the only way to collect the
animals without losing a lot of information is to preserve
the animal in place as you are collecting them. Often it involves
injecting the coral, for example, with a preservative and
then having to chisel or break that part of the colony free,
after which it is put into a labeled container, and more preservative
is added. Now, this all sounds easy enough, but then conceive
of doing it while floating weightless in the water column
with wave surge or currents tossing you around, all the while
trying not to destroy the adjacent animals or inject yourself
or your diving partner with the preservative, a potential
outcome that can really ruin your whole day. Then when you
get back to a laboratory, you are faced with the rather daunting,
and severely boring, task of microscopically examining the
gut cavities of all the preserved animals for food. Finally,
if perchance you do find some gut contents, you have to try
to recognize and count them. This may sound relatively easy,
but speaking from the experience of having to identify gut
contents, there are few tasks in biology harder than trying
to identify half digested parts of, for example, gelatinous
animals, or perhaps the feces of a fish that ate a gelatinous
animal. So… the bottom line is that data from corals
and many of the bottom-dwelling animals are few and far between.
|
|
|
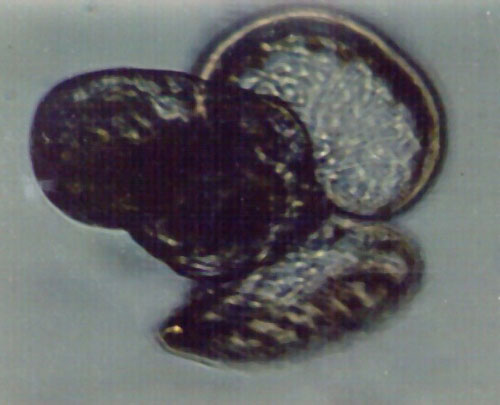
Figure
3. Other members of the "gelatinous zooplankton"
are larvae such as this small snail. The larval snail's
shell is to the left, and the two lobes of an extendable
feeding apparatus called the "velum" are extended
from the shell. These small larvae are both favored
foods of a lot of fishes, and surprisingly abundant
in the plankton. This animal was about 1/250th of an
inch across.
|
However, there are some data about what
coral reef fishes are eating, and those data may provide some
very useful insight in the feeding dynamics of this ecosystem
and our captive counterparts. One of the inherent difficulties
of the data that are available about feeding on a coral reef
is that the whole story is seldom told. For example, to be
able to assess how animals feed, one needs to know what is
available for them to feed upon. There are lots of data, most
of which are poorly taken, about what is available in the
plankton around a coral reef. And there are some data about
the feeding of coral reef animals on plankton. Unfortunately,
there are very few studies of what the animals are feeding
upon that are done with concurrent samples of the available
food.
One quite good study discussing zooplankton
availability and concurrent feeding by planktivorous reef
fishes has been published (Hamner, et al., 1988), and the
findings of that research are well-worth examining as they
provide one of the few records of the amount of food available
to reef animals. Additionally, and possibly more importantly,
the data shed some light on the fates of the various potential
foods that are moved over a reef crest by water currents.
These researchers examined a reef where
they were able to sample water flow from a depth of 25m (82.5
ft) up over the reef crest. The water flow through this area
was very nearly linear, so the water moved toward the reef
crest, swept upward and over the crest with little mixing
or turbulence. During a 12 hour period, 6,000 m3 (= 1,585,200
gallons or 132,100 gallons/hour) flows over that 1 m wide
portion of reef crest. This was estimated to bring 1,098,000
potential food items, about 70 percent of which are copepods
and larvacean tunicates. These items are the preferred prey
of the plankton-feeding fishes living on the reef front. Over
the space of that swath of reef front from 25m depth to the
crest, were about 500 fishes of 13 different species. Those
fishes consumed, over a 12 hour day, 1,180,000 items of food.
This was a wet weight of about 0.4 kg per day per meter of
reef front. This is roughly a pound of zooplankton food per
day. This all works out to 236 items of food per fish, per
12 hour period, or approximately 2.0 grams of food eaten per
fish per 12 hour period.
From these data (Hamner, et al., 1988),
it is apparent that during the daylight, most relatively large
(0.250 mm or roughly 0.01 inch diameter) zooplankton do not
make it to the reef at all. The authors note specifically
that the fish effectively feed constantly and the food transit
time in their guts is often short, sometimes only a few minutes.
These feces contain relatively large amounts of undigested
foods and are often eaten by other fish. Eventually, however,
a large amount of the zooplankton food that would have impinged
upon the reef does make it to the reef, albeit modified into
the form of fish feces. This is rapidly ingested by corals
and other benthic animals. At night, of course, most of the
zooplankton approaching the reef does encounter the benthic
reef animals. The number and volume of animals during the
night may be significantly higher than during the day as many
zooplankton animals will migrate to shallower depths during
the night, and these animals would be caught in the currents
and swept over the reef.
A couple of points are worth noting here.
The calculation of both feeding rates and plankton numbers
are approximate, and probably accurate within about plus or
minus ten percent. The fact that the number of potential food
items, and the number of eaten foods agrees as well as it
does, also within about plus or minus ten percent, is really
quite remarkable. Also, what is apparent is that the fish
eat ALL the plankton approaching the reef. NONE of it will
reach the reef during the day when the fish are feeding.
Additionally, the researchers looked at
the gut contents of 11 species of planktivorous fishes found
in their one meter wide swath. These species included three
species of fusiliers: Caesio cuning, Cuning's fusilier,
C. caerulaurea, the scissor tail fusilier and Pterocaesio
diagramma, the two-lined fusilier, as well as eight species
of damsel fishes: Chromis atripectoralis, the black
axil chromis, Neopomacentrus azysron, the yellowtail
demoiselle, Abudefduf whitleyi, Whitley's sergeant
fish, Amblyglyphidodon leucogaster, the whitebelly
damsel, A. curacao, the staghorn damsel, Pomacentrus
lepidogenys, the scaly-cheek damsel, Pomacentrus coelestis,
the neon damsel, and Pomacentrus molluccensis, the
lemon damsel. They examined the number of dietary items in
the guts of the fish, as well the percentage of fish which
had the items. The dietary categories they examined were:
algal fragments, copepods, larvaceans, salps, foraminiferans,
eggs, miscellaneous crustaceans, amphipods, gastropods, planulae
(from corals), chaetognaths, pteropods, and scales (from fish).
I modified and condensed these data and those data are summarized
in Figure 4, which shows the average number of fishes of a
given species containing the various dietary items. I condensed
the data from the two months, September and December, sampled
in the study, to give the single average values of Figure
4.
|
|
|
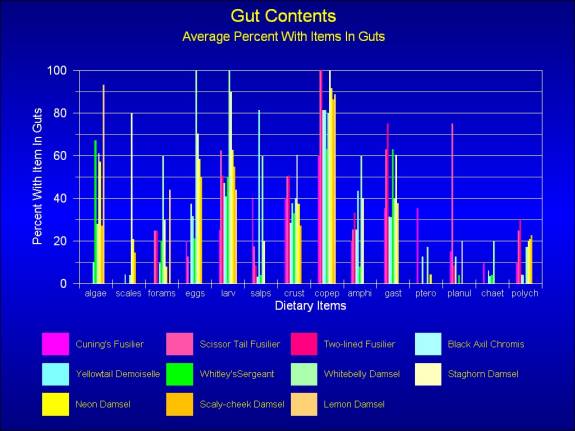
Figure
4. The average number of fish sampled of the given
species containing the various items as gut contents.
Data modified from September and December data, of Hamner,
et al. 1988. Fusiliers forage some distance from the
reef, and to differentiate them from the various damsels,
they are color coded with various shades of pink. The
various species of Pomacentrid (damsel) fishes are coded
with yellows, greens, and blues.
Click on image for larger version.
|
Some things are immediately apparent. All
of these fishes eat large amounts of crustacean prey, particularly
copepods. Larvacean tunicates were also a major source of
food, for all species, as were, to a lesser extent, pelagic
gastropods. There are some interesting, and obvious differences
in diets as well. Fusiliers do not eat algae, whereas all
of the Pomacentrids had some algae in their guts and some
had quite considerable amounts.
These data were the results of examining
the gut contents of 240 individual fishes, and the number
of prey items per fish was also enumerated. The number of
individual prey items in the gut contents was quite considerable.
The average number of food items in the guts of black-axil
Chromis in December was 665, and for the average yellow tail
demoiselle had 1,036 during the same sampling period. The
average number of copepods in the guts varied from two to
over 200, and the fusiliers typically had far more than the
pomacentrids. The diversity of dietary items per fish species
was generally high with most species eating at least items
from ten of the 13 enumerated food categories.
Fish Diets
From this study, it is apparent that these
fish are feeding continuously throughout the daylight hours.
They are eating small items, but on the average they eat an
item of food every three minutes all day during a twelve hour
day. During that period they eat an average of two grams of
food per day. As a comparison, during my food and additive
study (Shimek, 2001) Ocean Nutrition Products, such as Formula
1, had 70 cubes per 7 ounce package of food. That meant each
cube weighed 2.8 g. On the average, if you wish your fish
to have the same mass of food that they are likely to eat
in nature, presuming the data of Hamner et al., 1988, is applicable
to other fishes, you should feed each fish in your aquarium
that is the average size of a damsel fish, the equivalent
of about 70% of a cube of this food per day. Large fishes
would get proportionally more.
Coral Reef Invertebrate Diets
During the day on a natural reef, it appears
that virtually no moderately large zooplankter would reach
the coral on the reef's face. Nonetheless, this area would
be bathed in a diffuse rain of particulate organic material
derived from fish feces, dissolved material and microzooplankton.
Much of this is generated in the habitat immediately adjacent
to the reef by the action of the planktivorous fishes, but
a lot of it comes from more distant regions. A lot of it consists
of remnants of larvacean tunicate houses, (Alldredge, 1972),
but it has other components as well (Alldredge and Silver
1988). Larvacean tunicate houses are gelatinous or mucoid
constructions comprised primarily of the mucoid protein, chondroitin
sulfate, and embedded or covered with coccolithophores, cyanobacteria
and occasionally diatoms. Additionally, of course there are
fish and copepod feces and other materials of rather dubious
parentage and food value.
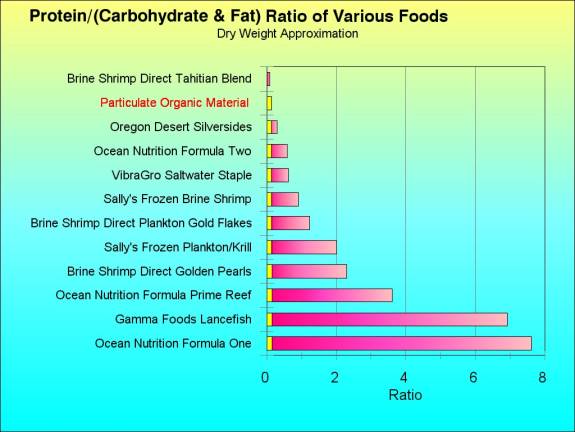
The particulate organic material that
reaches the reef face proper to be ingested by reef animals
in the daytime is not particularly high quality food. The
average dry weight Carbon to Nitrogen ratio of the particulate
material is about 7.8:1, which is equivalent to the Nitrogen/Carbon
proportion of 0.127 (Alldredge and Silver, 1988). This is
a food high in carbohydrates and low in proteins. Such a food
ratio is very different in content from the average aquarium
food, Figure 5. If the weight of protein in a food is divided
by the weight of the carbohydrates and hydrocarbons in a food,
a rough estimate of Nitrogen/Carbon may be obtained, and I
did this manipulation for some of the foods from my earlier
study of foods and additives. This estimate of Nitrogen to
Carbon is likely to be a somewhat too high as there is carbon
in the proteins. Additionally, the presence of oxygen and
hydrogen would make the results a bit fuzzy, but these latter
elements may largely cancel themselves out on both sides of
the ratio. Aquarium food is much higher in nitrogen to carbon
than is the typical food item that hits the reef during daylight
hours. During the night, the quality of food impacting on
the reef animals may be very much higher, consisting of the
actual (" = unpreprocessed") zooplankton.
The amount of protein in virtually all
aquarium foods is well in excess of what is found in natural
reef foods (Figure 5). In the calculations used to construct
Figure 5, the moisture contents of the various foods were
removed, so these are data compared on a dry weight basis.
Most of the wet or frozen foods listed are 75 percent to 85
percent water, so to compare them with regard to weight of
the normal sample, see Shimek, 2001. It is apparent from Figure
5 that most marine aquarium foods are very highly enriched
in protein with regard to natural Particulate Organic Material.
|
|
|
Figure
5. The amount of Protein divided by the sum the
Carbohydrates and Fats from some common aquarium foods
(Data from Shimek, 2001); based on a dry weight approximation.
The value for natural particulate organic material is
from Alldredge and Silver (1988) and is indicated on
all foods below it.
|
Marine Aquarium Ecosystems Foods Differ Significantly
From Natural Ones
Several major differences between marine
aquaria and natural reef ecosystems with regard to food should
be apparent. In this analysis, I am largely ignoring the bacterial
component of the foods. There is not much an aquarist can
do about this component of the food one way or another. However,
all aquarists may significantly control the amount of particulate
food in their aquarium. This food will mimic either the zooplankton
or the particulate organic material components of coral reef
feeding dynamics. For the animals in a system to be healthy,
those animals must be fed foods that more-or-less duplicate
the qualities of their natural foods, and they must be fed
in a more-or-less normal matter. Reef aquarium foods and feeding
regimes tend to fail rather spectacularly on both accounts.
The standard reef aquarium is probably
fed once about once a day (Shimek, 2002), and the average
daily feeding ration weighs 15.39 ± 15.90 grams, or
roughly a half of an ounce, wet weight, of food. On a natural
reef, this would be enough to provide roughly eight damsel
fish with their normal daily allotment of food. Unfortunately,
this amount of food all occurs effectively at once or over
a very short period in an aquarium whereas on a natural reef
it would occur over a 12 hour period. Additionally, aquarium
food is a relatively high-protein material. When most reef
fishes encounter planktonic patches of food, they eat voraciously,
and material gets passed through their guts in a rapid manner
resulting in incomplete digestion. This is precisely what
happens to many fish in an aquarium when it is fed. If you
watch some of your plankton feeding fishes, such as clown
fish or damsels, you will see that shortly after the initiation
of feeding they start defecating food at an increased rate.
In effect, they are pumping food through their guts. The faster
the passage of the food through the gut, the less the fish
get from it. Perhaps in nature this doesn't matter, as the
food is always coming at them. In the aquarium, this effect
could be quite deleterious.
In aquaria, fish that naturally feed consistently
on small particulate material throughout the day are being
forced to exist on bulk feedings once a day or with less frequency.
Under such conditions, the animal is going through continuous
cycles of near starvation followed by satiation followed by
near starvation… This cyclic feeding simply must have
a deleterious effect on the fish. Under such situations one
could expect lower than normal growth rates, higher stress,
increased susceptibility to disease and possibly problems
with nitrogen metabolism. During the periods of low food availability
the fish would potentially metabolize excessive amounts of
protein, resulting in excessive ammonia production. Something
similar would be seen with the sessile animals on the rocks
of a reef aquarium. Here the food intake is likely intensively
periodic with significant periods of non-nutrient input. Growth
will also likely be reduced and the animals stressed.
The amount of food impacting on the reef
over the course of a day is substantial. Over a section of
a natural reef about three feet on side, flows a continuous
flood of water carrying with it about 2,000,000 food items
with an aggregate weight of about two pounds in a 24 hour
period. These tiny food items would be like a rain of diffuse
nutrition on the reef and reef animals, particularly the fish.
Suggestions
It is apparent that coral reef planktivorous
fishes, and this is most of those kept in aquaria, would benefit
from changes to the normal aquarium feeding regimen.
|
•
|
They should be fed by some sort
of continuous feeding apparatus. |
|
•
|
The food dispensed by such an apparatus
should be particulate in nature, and very small. The largest
sizes should probably be on the size of a brine shrimp
or smaller. |
|
•
|
Such food need not be specifically formulated to be
highly nutritious: rather it should be of low to moderate
nutritional value. If aquarium fish are able to eat more
continuously and slowly, they will get much more nutrition
out of each food item than they do now. Feeding a low
quality food should result in significantly less nutrient
accumulation than is now commonly seen in tanks. |
In effect, we need to turn our feeding
regime on its head. Rather than feeding a small amount of
highly nutritious food once a day, we should be feeding a
large amount of low nutrient value food frequently. Such a
feeding regime as this should reduce significantly the amount
of pollution effects in reef aquaria. Additionally, there
would not be a daily pulse of nutrients to temporarily overwhelm
the biological filter. In turn, there would less potential
growth of problem algae and the development of a more balanced
and easily controlled assemblage of animals within the tank.
|