|
Introduction:
Regular readers
of this column will be aware that I have a fascination with
the bizarre and weird. I guess this comes from growing up
in central Montana. If one knows where and what to look for,
some of this area may be seen to be badly disturbed ecologically;
however, in comparison with most of the United States, much
of it looks absolutely pristine. In such an area wildlife
abounds, and when I was but a mere lad I became familiar with
all of the common sorts of the animals found in the area,
from horses to horsehair worms to toads, horned and otherwise,
various bugs, Black Widow spiders, and fishes of all sorts.
Once I become really familiar with an animal or animal type,
I "file it away" and go on to something different.
Lacking a microscope when I grew up, I didn't learn about
microscopic animals until I was in high school, and then only
cursorily. It wasn't until I reached college that I began
to explore the wonderful world of the miniature animals and
other organisms found all around us (and in us, as well, but
that is another story).
Although my introduction to these, the smallest animals,
came with the examination of freshwater microscopic organisms,
similar organisms are common in marine systems. In fact, with
few exceptions, most of the groups containing small animals
are more common in marine ecosystems than in freshwater ones.
That having been said, the animal group of focus in this column,
the rotifers, is somewhat abnormal in that it is more diverse
in freshwater environments than in salty ones. Subsequent
columns in this short series on small animals will discuss
animals that reach their acme of diversity in marine ecosystems,
but that is not the case with the rotifers. Although rotifers
may be easily found in salty water, they are both more diverse
and often more abundant in freshwater habitats.
When discussing small organisms, the discrimination of what
is, and what isn't, an animal becomes rather blurred. Some
of this blurriness has come about in the last half century
or so, directly as a result of research that increased our
knowledge base. When you have only a couple of data types,
it is easy to use them to categorize things and put them into
some sort of order. When the amount and kinds of data become
diverse, such categorization may become very difficult. Virtually
all of the early examination of microscopic organisms used
light microscopy as its primary tool. This is an excellent
way to examine these creatures, but as with all ways of observing
something, it imparts a bias. Basically, the bias is a simple
one: "If the creatures look alike, they probably are
alike." This seems simple and intuitively correct. For
many large organisms this bias works pretty well. However,
when examining the "teeny-tiny," such an approach
becomes unworkable. Its major problem is one of structural
complexity, or more correctly the lack of structural complexity.
For example, while large animals are almost always structurally
complex, having organ-systems, many types of tissues and cells,
literally by the trillion, smaller organisms are often much
simpler in structure. If aspects of that structural complexity
are used to categorize or "pigeon-hole" organisms,
then what happens to the ability to categorize when the complexity
isn't there? When observing the apparent simplicity of small
organisms, the question arises, "Are these organisms
truly fundamentally simple or have they become simple while
evolving from much more complicated ancestors?"
Life in the small lane is constrained by all sorts of odd
factors, and as natural selection has worked in these environments
just as well as in the realm of the large and ponderous, the
question becomes one of convergences. In other words, "Do
the structures we see in two different organisms reflect a
common ancestry (divergent evolution), or do the organisms
only seem similar due to superficial similarities caused by
convergent evolution resulting in the loss of complex structures
that yields similar simple appearances?" This fundamental
question remained largely unresolved until the mid-1980s when
examination of the actual genome
started to become commonplace.
As a result of such studies, the whole "world-view"
of how living organisms are related - and grouped together
- has changed, and those changes continue unabated. The classification
schemes and presumed relationships between groups that I learned
and taught as recently as five years ago have been shown to
be based upon inadequate data and have had to be revised.
The species haven't evolved much in that period, but our understanding
of their relationships has been altered by the inclusion of
a lot of new, supportive data. This has allowed fine-tuning
and precision within the "Tree
of Life" that hitherto had been only surmised.
If we accept the premise that organisms with similar genomes
are related, then their degree of relatedness can be determined
by measuring the degree of difference between the genomes.
Degrees of relatedness determined by genetic testing can then
be compared with degrees of relatedness determined by other,
more traditional methods, such as skeletal or structural analyses.
With most of the larger animals, there have been few surprises;
generally, the data sets concur pretty well, and although
there have been a number of "revolutions" in the
relatedness of major animal lineages, generally what had been
thought to be closely-related groups usually turn out to be
so. This hasn't been the case with many of the smaller organisms.
Because of all these changes, the old dichotomy of life as
belonging to either the animal or plant kingdoms has been
discarded.
Presently, the classification scheme used to categorize organisms,
based on genetic and cellular features, is one whose largest,
most inclusive groupings are called Domains of Life. Generally,
there are considered to be three
Domains. Within, or under, each Domain are subdivisions
called Kingdoms. The animal and plant kingdoms have been redefined,
and both are now regarded as containing only multicellular
organisms. Under the present classification scheme may be
separate kingdoms for many of the major algal groups as well
as for many unicellular organisms previously categorized as
protozoans; however, that sort of standardization is in the
future. For the purposes of this essay, though, the important
point to remember is that all animals are composed of more
than one cell.
Multicellularity
Animals, then, by definition, have
more than one cell, which fundamentally limits organisms'
possible "smallness;" animals can get very large,
but they can't become infinitely minute. If a body is to be
made of functional components called "cells" and
that body is constrained to be small, then there is an absolute
limit below which an organism made of multiple cells cannot
be made. Below that limit, each cell would simply not have
enough volume to contain what it needs to function in the
context of a multicellular organism. Organisms constructed
without cells, or organisms constructed of only a single cell,
may be much smaller, but for organisms made of multiple cells,
there is a lower size limit. The smallest common animals are
the subject of this essay; these are the strange animals called
"rotifers." Only the very odd animals called "Loriciferates"
are smaller than rotifers, but loriciferates are so poorly
known - no living specimen has ever been studied - and so
seldom seen that it is hard to discuss them. Rotifers, on
the other hand, are exceptionally common and very well known.
Although small (large rotifers seldom exceed a fiftieth of
an inch in length and some of the smaller ones are only about
40 µm, or about 0.0016 inch, long), they are among the
most ubiquitous freshwater creatures, and many species are
common in marine areas as well. When examining rotifers, it
is astounding to realize that these complex animals, containing,
for example, several organ systems, are smaller even than
many single-celled organisms such as Paramecium and
Amoeba that used to be called protozoans.
The animals normally referred to as rotifers share a single
unified body plan and are readily recognized by anybody who
has had the opportunity to see one. Even so, they are relatively
diverse; about 2200 species are described. Taxonomically,
they are placed into the Phylum Rotifera, which is subdivided
into several major subgroupings and a couple of smaller groups.
The recent "fly in the taxonomic ointment" is, however,
that what had hitherto been considered a discrete phylum,
or major group, of parasitic organisms, the Acanthocephala,
are descended from, and should be considered as, highly modified
rotifers. Genetic research has determined this is unequivocal;
the Acanthocephalans
are modified rotifers, but just how, exactly, they fit
within the so-called phylum Rotifera is open to question and
has not been resolved. The resolution of that question, among
a number of other questions, renders meaningless the discussion
of the relationships between major subgroups in rotifer taxonomy.
However, the purely "classical" rotifer groups themselves
haven't changed much, and so may be discussed without loss
of information.
Adult and juvenile rotifers have no way to avoid desiccation
and must therefore live in water. Freshwater rotifers often
seem to be specialized to live in specific transitory environments,
and part of that specialization has been the development of
some water-resistant reproductive stages. These are basically
eggs with highly resistant shells. Rotifers may make their
homes in some habitats, such as the fluid captured deep inside
a tubular flower blossom, or the muck of a simple mud puddle,
that would seem unlikely places to be dignified with the name
"ecosystem"; nevertheless, these small transient
bodies of water have all the attributes of much larger ecosystems:
a physical environment, biota and food webs distributing and
transferring energy and materials. And in them, rotifers thrive.
In point of fact, these small "here today, gone tomorrow"
habitats are specifically the places where rotifers are most
abundant and diverse, and many of their attributes appear
to have been adapted for maximal utilization of such environments.
Their small size notwithstanding, some species of rotifers
are herbivores, others are carnivores, still others are suspension-feeders
and yet others are parasites.
Morphology
Rotifers are sometimes described
as elongated worms, but they really don't look very wormy.
Their body
is often divided into three regions, which often "telescope"
in and out of one another. Their body is covered in a rigid
cuticle that holds its shape very well, so unlike worms which
twist and turn and appear to be almost infinitely flexible,
rotifers' shapes are relatively constrained and consistent.
Many of them look like self-mobile miniature torpedoes. One
of the rotifers' best identifying characteristics is that
the front of the animal bears a crown of large, evident, beating
cilia. These cilia whip, flex and bend, and in the process
they move water past themselves or they move the animal through
the water (see some rotifer movies: 1,
2,
3).
Rotiferan cilia function to move the animals through water
and additionally, they bring food particles to the
animal. Rotiferan cilia are arranged in circular patterns
to form the "corona," or ciliary crown, on the front
of the animal. When these distinctive cilia are actively beating,
they tend to look like moving wheels, so much so that for
many years, rotifers had, among biologists, the peculiar name
of "wheel animacules." In fact, the name "rotifer"
is derived from a combination of the Latin roots "rota"
or "rotula," meaning "wheel," and
the Greek root "phoreus," meaning "a
bearer." In other words, they are animals that bear wheels.
|
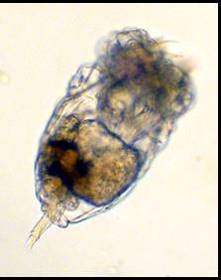
Figure 1. A couple of marine rotifers. See Figure
3 for anatomical structures. Both animals are between
0.05 and 0.1 mm (0.002 and 0.004 in.) long.
|
|
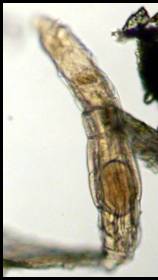
Figure 2. Not all rotifers are mobile swimmers.
This individual of a sessile species was living fastened
to a hydroid stalk (the brown structure running vertically
through the image). Sessile rotifers often use their
lorica as a "house" to withdraw. The corona's
cilia are visible as thin, straight lines in the image.
This animal was about 0.05 mm (0.002 in.) long.
|
Behind the crown of cilia, the majority of the body is called
"the trunk" and is covered with a hard shell, often
called a "lorica." This lorica is transparent and
appears to be the body wall; it is not distinguishable by
color or shape. It is often composed of cylinders or rings,
and may give the appearance of being segmented. Many rotifers'
bodies are "telescoped" with one section of the
trunk sliding into another. The lorica is a peculiar and complex
structure; its external part is formed outside the animal
by secretions of the body surface. This part is non-living
and made of proteins. There also is an internal portion of
the shell formed of cellulose-like materials. Consequently,
in some regards, a rotifer's shell is somewhat like, but on
a much smaller scale than, a turtle's bony shell, the inside
of which is living, and the outside, which may be dead.
The last part of the rotifer's body consists of the foot,
which often has two lateral branches called toes. This foot
is used in locomotion, but not as an ambulatory organ. Rotifers
usually fasten to the substrate using their foot to provide
adhesive purchase. The foot contains duo-gland adhesive structures,
so when the animal wants to stay in one place, it touches
its toes to the substrate and secretes the adhesive. When
it wants to move, the toes secrete a second substance that
releases the toes from the glue, and the animal swims off.
Many rotifers creep along the substrate almost in an inch-worm
manner using their toes and adhesive glands to fasten to the
substrate as they crawl along.
|
Figure 3. The basic structures found in a rotifer,
using the common aquarium food species, Brachionus,
as an example.
|
Rotifers are adapted to many different modes of life and,
not surprisingly, these are reflected in their bodies' different
shapes and functionality. The swimming forms generally are
rigid and many have spines to protect them from predators.
Often, these spines are variable in size and extent. In some
species of Brachionus spines appear only when its major
predator, another rotifer called Asplanchna, is present.
For its size, Asplanchna
has a huge mouth and eats Brachionus individuals by
engulfing them whole. When Asplanchna are absent, Brachionus
is spineless or has, at best, very short spines. If Asplanchna
appears, the
longer spines are found in the NEXT generation
of Brachionus. The presence in the water of specific
chemicals, small protein fragments, released by Asplanchna
as it swims around, causes the production
of spines in Brachionus embryos. These spines can
be large enough to make the prey inedible; the predator simply
can't fit the prey into its mouth.
Rotifers are very peculiar animals, however, much more so
than people who have examined them only in passing ever realize.
Most people who have taken any biology or even general science
courses know that "ALL" living things are composed
of cells. Unfortunately, I suppose, no rotifer has been to
these classes, and consequently they don't know that they
are supposed to be made of cells. So... they aren't. The majority
of rotifer structures and tissues appear to be made of standard
cellular tissues, but closer examination shows that most of
them lack cell membranes that completely delineate those cells.
In effect, each "tissue" or "organ" is
a blob of protoplasm containing several nuclei or cellular
control centers. Unlike most animals, however, these nuclei
are not found in cells, but float together in their little
"blobs of goo." These protoplasmic masses, which
have many nuclei but lack cellular membranes separating them,
are called syncytia. Syncytia seem to be characteristic of
many smaller animals such as the rotifers and nematodes, in
addition to some other odd groups such as the glass sponges
(Hexactinellids).
In the smaller animals where they are found, they may simply
be a biological solution to microminiaturization. Check out
Coles Catalogue
for the specials of the week.On this
scale of animal dynamics, expending the energy and materials
necessary to make membranes to delineate cells may not be
cost efficient. In effect, the animal is so small that the
benefits of being cellular, presumably a compartmentalization
of structure and function, don't outweigh the costs of producing
the cells or the costs of moving materials across cell membranes.
Rotifers and a few other groups of animals, including nematodes,
share another odd property. It is called "Eutely,"
or "cellular constancy." Eutely means that all adult
members of a given species of animal have exactly the same
number of cellular nuclei in exactly the same place. For example,
in the common freshwater rotifer Epiphanes,
their 959 nuclei are arranged so that 172 are in the corona,
108 in the trunk and foot epithelium, 19 in the pedal glands,
22 in the circular muscles of the body wall, 40 in the coronal
retractor muscles, 183 in the brain, 68 in the peripheral
nervous system, 167 in the jaw apparatus called the mastax,
15 in the esophagus, 39 in the stomach, 12 in the gastric
glands, 14 in the intestine and 25 others distributed in other
places. Every Epiphanes will always have this number
of nuclei in exactly the same place in its body.
Such a stereotypical body plan is unlike what is found throughout
most of the animal kingdom, and it has some profound consequences
for all animals that possess it. On the down side, because
no cell, or, more correctly in this case, any nucleus, has
any latitude in its genetically determined position or function,
there can be NO repair of injuries. Simply put, a damaged
rotifer dies. The trade-off is that such a deterministic body
plan apparently allows very rapid growth and development.
Many rotifers live fast, die young and leave a little blobby
corpse. Many of them have a life span of only about a week.
Numerous species may reach adult size less than a day after
hatching from the egg. Such rapid development may be possible
only with a "slimmed-down" genetic complement. In
effect, many of them are lean, mean, reproductive machines.
Once an egg reaches a puddle it can hatch and be ready to
reproduce within a day, and several generations may be found
in a mud puddle that lasts a week. All of this appears to
be the result of countless generations of rotifers adapting
to transient habitats by reducing their genetic code to the
barest necessity.
All of these genetic reductions might tempt an observer to
think that they have reduced other things, such as organ systems,
as well. This definitely does NOT appear be the case. For
example, their nervous system is remarkably well developed.
Consider that the Epiphanes mentioned above has 183
nuclei in its brain. This means that 183/959, or 19.1%, of
the animal's nuclei and protoplasmic material is devoted to
the brain. In comparison, an adult human's brain contains
no more than about 2-3% of the body's mass and cellular structures.
On this scale, at least, rotifers are proportionally and significantly
more "brainy" than most politicians, if not the
rest of humanity. Rotifers have a well-developed, bilobed
brain. Additional components of their nervous system include
several sensory tentacles, used to detect chemical and tactile
stimuli. They generally have at least one photoreceptor, commonly
called an "eye," and possess tactile cilia as well.
The rotifers' muscular system is well-developed, with individual
cells working as independent, separate muscles. Probably as
a result of their small size, they lack the muscles comprised
of many cells and the muscle bands found in other, larger
animals. Nonetheless, they are capable of a wide array of
muscular movement and precision.
|
Figure 4. A marine rotifer, with the position
of its muscles indicated by the red bands on the right
image. The animal was about 0.1 mm (0.004 in.) long.
|
Their digestive system is complex, consisting of three major
regions: a foregut, a midgut and a hindgut. The foregut consists
of the mouth and the food grinding apparatus, or mastax. The
mastax is a large (for a rotifer) muscular mass surrounding
an inner mouth region. In it are three grinding jaws arranged
so that its gut in cross-section has the appearance of a pie
made of three slices, but with each slice being a massive
grinding plate complete with ridges and teeth. The short esophagus
passes the food to the large stomach, which is surrounded
by a digestive gland. Digestion, unlike the condition found
in many invertebrates, is extracellular and occurs in the
stomach cavity. Nutrients are absorbed through the stomach
lining and transferred directly to the digestive gland by
syncytium-to-syncytium transport. Food is further processed
in the digestive gland, in a manner probably analogous to
what occurs in a vertebrate's liver. The short intestine also
absorbs some nutrients but functions mostly to transfer food
remains to the anus.
Nitrogenous waste, mostly ammonia, is excreted by the kidneys
through a flame bulb, or protonephridial
construction. Excess water is also moved out of the body
though the same tubules, flushing the system. Rotifers are
so small that they have no need for a circulatory system.
Nutrients are simply passed into the fluids filling the body,
and moved from place to place by the movement of the whole
animal.
As with many of the rotifers' other characteristics, their
reproduction is an exercise in strangeness. In one of the
groups, the "Bdelloid
rotifers" commonly found in freshwater, no males
have ever been seen and likely do not exist. The females in
this group produce parthenogenic
eggs. These eggs develop into small juveniles before they
are released from their parent, and actually are clones of
their mother. To further maximize reproductive potential in
some species, when the juvenile exits the mother, it already
contains developing embryos. In a very real sense, they are
truly born pregnant.
One of the tenets of biological science over the last 60
or so years is the concept
of the biological species. Proposed by Ernst Mayr in 1942,
a "biological" species is supposed to be a group
of animals, all of which can interbreed with each other. Mayr
erected the idea of the biological species in response to
the "morphological" species, or "morphospecies,"
concept, wherein a species were all of the animals that looked
alike. Of course, the problem with the morphological species
is that "looking alike" is a lot like the concept
of beauty. How "alike" animals are varies with their
beholder. The morphospecies concept can create all sorts of
problems depending upon who is deciding what is "alike."
One biologist, C.
Hart Merriam, over a period of years described more 90
species of different Grizzly bears. We now recognize that
all of these so-called species were, in fact, just habitat
or even individual variations. Such an absurd subdivision
of any group creates a nomenclatural fog that obscures and
retards actual research. Consequently, Mayr's attempt to create
biological species was considered to be a great leap forward
in biological thinking. However, it didn't take into account
such organisms as bdelloid rotifers. In the biological species
concept as used by Mayr, either no bdelloid rotifer group
constitutes a species, or each bdelloid lineage is
a species unto itself. To some extent, such a situation is
simply a matter of nomenclature. On the other hand, it also
indicates that these animals are very different from sexually
reproducing animals, including other rotifers. Having no sexual
reproduction and no means of expressing variability, bdelloid
rotifers may have, in effect, opted out of evolution. They
don't appear to change or to have any adaptive capability.
On the other hand, they are widespread and perfectly adapted
to habitats that will probably always be present, at least
for a long time, evolutionarily speaking. Other rotifer types
do have males, at least part of the time, and by examining
their life history patterns, it is easy to see how the parthenogenesis
of bdelloids got started. Once it occurred, that lineage was
locked into a form that never can vary.
The sexually reproducing rotifers constitute the other, and
largest, group of rotifers. These animals, including the aquarium
food genus, Brachionus and other cultured rotifers,
consist of separate sexes that do not look alike. The small
males inject sperm through the female's cuticle. The female
typically has a small ovary which contains about 20 nuclei
(eggs). These particular species' life cycle is cued to environmental
conditions. As long as the animals are growing in good conditions,
thin-shelled eggs are produced that have two sets of chromosomes.
These eggs are produced parthenogenically. When the environmental
conditions in the habitat turn bad, if the animals are overly
crowded for example, a different kind of egg is produced,
with only one set of chromosomes. If the female doesn't mate,
the egg is unfertilized and develops into a male. If she mates,
it gets fertilized and develops a thick, resistant shell.
When this occurs, it is called a "resting egg."
It will not hatch until environmental conditions improve.
This type of life cycle is tailor made for aquarist cultures.
The resting eggs may be sold as starter "cysts"
or "eggs." Once rehydrated in a good culture medium,
the animals rapidly grow and produce a lot of offspring. As
long as the culture conditions remain good, that is, as long
as there is no accumulation of waste materials, the oxygen
concentration remains high, and steady, carbon dioxide remains
low, and there is plenty of food, the animals continually
reproduce, and can be harvested and fed to the animals in
the reef tank. Once the culture conditions deteriorate and
the population starts to crash, resting eggs are produced
and , with care, may be collected, dried and used to start
a new culture.
Rotifers make a good food for many small fishes, fish larvae,
or invertebrates specialized on small planktonic animals.
The major drawback of these cultures is that although the
rotifers used in them, mostly one species or another of Brachionus,
may tolerate saltwater for short periods, they really don't
do well in it, and therefore don't last in the tank for any
appreciable period of time. Nonetheless, they are easy to
grow and do make a good transient food source that mimics
the small gelatinous zooplankton that many animals feed upon.
Additionally, they are harmless to anything in the tank and
will not spread diseases.
Truly marine rotifers do exist, but they are somewhat uncommon
and hard to culture, and most of them are not planktonic.
One group is even wholly parasitic on crustaceans. Nevertheless,
the available rotifer cultures do make a good substitute for
the small gelatinous zooplankton lacking in many of our systems.
|


)
)

