|
A Stealth Topic
Out of all the topics
far from the minds of most reef aquarists, it would probably
be difficult to find one further from consciousness than the
consideration of how the animals in their tanks eliminate
waste materials. If this is even considered at all, it is
thought of along the lines of, "The animal poops it out,
and from then on the scavengers/detritivores get rid of it."
This is, of course, a very concise way of thinking about the
elimination of uneaten food from the digestive tract. Unfortunately,
it has nothing at all to do with what biologists consider
to be waste. Not to put too fine a point on it, but fecal
matter is nothing more than uneaten, partially digested and
processed food. Although relatively unsightly, and probably
a bit messy, this stuff is generally not overtly poisonous.
In fact, many reef animals, among them many popular reef aquarium
fishes, are coprophagic. "Coprophagic" is the jargon
term for what might politely be termed, "excrement-eating."
Ah… Euphemisms are alive and well; I long for the directness
of the old Angles and Saxons. However, the point is obvious;
if a lot of animals eat the stuff, it isn't toxic. It is preprocessed
food.
Actual waste materials are something else altogether. Strictly
speaking, to a biologist, only a couple types of materials
are truly waste materials. These are the byproducts of cellular
respiration and protein metabolism, which in most animals,
are carbon dioxide and ammonia, respectively. Carbon dioxide
removal is governed by many of the same rules that dictate
how ammonia is removed, but carbon dioxide is much less toxic
than ammonia, and often its removal is coupled with oxygen
uptake in animals' respiratory systems, which I will discuss
in a future column. Ammonia removal is somewhat more complicated
and is the topic of this column.
Amino Acids
Proteins have been called "the
building blocks" of animals. They are large molecules
comprised of subunits called amino acids. Around 20 amino
acids are found commonly in nature, and well over 100 are
less common. Amino acids are small molecules with a relatively
straightforward basic structure. At one end of the molecule
is an organic acid group. An acid is simply a molecule that
releases a proton, or hydrogen ion (H+),
in a solution. The most common organic acids contain a -COOH
group, which ionizes in water to become -COO-
+ H+. Vinegar is one of
the simplest organic acids, and it can be represented by the
formula, CH3COOH. Adding
one -CH2 group to vinegar
gives CH3CH2COOH,
which has the common name of proprionic or propanoic acid.
The basic backbone of an amino is a proprionic acid molecule
with an amine, or -NH2,
group substituted for one of the hydrogen atoms on the middle
carbon atom, giving the formula CH3HCNH2COOH.
This is the formula for one of the simplest amino acids, alanine,
and it has the typical amino acid structure, which is shown
below.
|
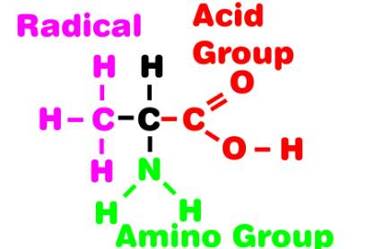
Figure 1. The diagrammatic generalized structure
of alanine, a simple amino acid. All amino acids have
a similar basic structure possessing the acid and the
amino groups; only the radical group differs between
different amino acids.
|
Proteins are assembled in a cell by chemically bonding a
great many amino acids, with the structure and properties
of all proteins ultimately determined by their amino acid
sequence and how these molecules are folded into complex shapes.
In living organisms, all larger molecules have a finite "lifetime,"
after which they are disposed of. When proteins are broken
down, enzymes slice them back down into their component amino
acids. These amino acids may or may not be further broken
down and their constituents harvested for new uses in the
cell. If they are broken down completely to their component
parts, the critical part of each amino acid is the amino group.
All other parts of amino acids can be recycled and reused
by animals, but the amino groups cannot be disassembled into
nitrogen and hydrogen atoms; they remain together, and therein
is the problem.
Ammonium Toxicity
During normal cellular metabolism,
the various chemical constituents are constantly being recycled
by breaking them down into their component parts and then
reassembling them into something useful. However, amino groups
are a dead-end in this regard. Unlike virtually all other
chemical bonds, animals cannot break the nitrogen-to-hydrogen
bond, and when the amino group is separated from any molecule
in an animal during a catabolic
reaction it immediately forms the exceedingly reactive
ammonium ion, NH4+.
This ion is easily capable of combining with, and destroying,
many vital and necessary cellular chemicals under the conditions
found in cells. This extreme reactivity makes ammonium ions
exceedingly toxic to many organisms, even in very low concentrations.
Obviously, this toxicity is the major reason that aquarists
monitor and attempt to minimize dissolved ammonia concentrations
in their tanks. Dissolved ammonia gas is simply ammonium ion
in water.
Ammonium ion is no less toxic to a single cell than it is
to the whole aquarium, and the need to keep its concentration
low is one of the driving forces of natural selection in many
animal groups. The removal of ammonium ions from an organism
is what biologists mean when they speak of waste elimination,
or excretion, from living organisms.
|
Figure 2. The tissue layers of corals and sea
anemones are so thin that any ammonia derived from the
digestion and subsequent utilization of the protein
keratin in this feather will diffuse out of this anemone,
directly from the epithelial cells that comprise its
body, into the surrounding water.
|
Size Related Solutions
Excessive ammonium is no problem for
the smallest animals, and for those acellular
organisms commonly referred to as "protozoa." These
organisms are so small that the ammonium ions simply diffuse
out of the cell. Diffusion works in response to a concentration
gradient, and as long as the ammonium concentration is lower
outside the cell than in it, ammonium will diffuse out of
the cell. This process is very efficient at moving materials
in solution, but it works best over very small distances.
At distances greater than a few small parts of a millimeter,
diffusion rates begin to drop off and rapidly become too slow
to prevent a rapid, and deadly, accumulation of ammonium ions.
|
Figure 3. Small acellular organisms, such as
this small planktonic ciliated protozoan about 0.1 mm
long, have no special excretory systems. Waste ammonia
simply diffuses out of the organism over the body's
surface.
|
The ammonium accumulation that occurs inside cells that are
rapidly metabolizing is one of the factors limiting cells'
maximum sizes. It is also of profound significance to aquarists.
Many aquarium animals lack any specialized organs or structures
for removing nitrogenous wastes such as ammonium ions. Although
it might seem counterintuitive, this is true of many larger
organisms as well as smaller ones. Most large and complicated
animals have very efficient kidneys and can regulate nitrogenous
wastes quite well, but many of the larger aquarium organisms,
such as corals and sea anemones, have a very simple structure
containing no excretory organs at all; nonetheless, they still
mange to attain, apparently, a quite large size without suffering
the deleterious effects of ammonium poisoning. The answer
to this paradox, of course, is that looks are deceiving. While
many corals appear to be large animals, their volume of living
tissue relative to their entire size is quite small. Most
corals are, essentially, very thin living tissue sheets layered
onto a massive, non-living, calcareous skeleton. Such animals
depend on their high surface area-to-volume ratio to provide
enough surface area for the diffusion of materials such as
ammonia and dissolved gases. Although it appears that sea
anemones are large lumps of living matter, as a practical
matter they also have a high ratio of surface area to volume.
Most of the mass of a sea anemone is made of non-living, organic
materials such as collagen and other fibrous materials located
between the thin outer and inner living tissue layers.
Activity Forces Changes
Actively moving animals have higher
metabolic rates than do sessile ones, and this means that
animals that move are particularly size limited by their surface
area-to-volume ratio. Unless they have some efficient way
of performing gas exchange and removing poisons from their
systems, they have to remain small. The bodily shape of the
larger flatworms is thought to be a response to competing
naturally selective pressures. These animals are essentially
two-dimensional; although they may be quite long or wide,
their thickness may approximate that of a piece of paper.
There are a great many reasons why it is good for animals
to be large. However, there are some significant problems
to overcome before large size is attainable. One of the primary
hurdles appears to be the development of a functional kidney
and excretory system.
The largest free-living and active flatworms are about five
feet (1.6 m) long, one inch (2.5 cm) wide and the thickness
of a couple pieces of paper. These animals, like most of the
larger flatworms, have an excretory system consisting of numerous
internal structures variously called flame bulbs, flame cells,
solenocytes or protonephridia. All of the names refer to similar
structures. However, they were initially seen and described
from different animals and were named independently by different
biologists.
|
|
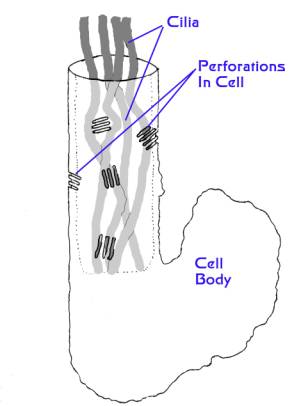
Figure 4. A diagram of a flame bulb cell.
The top of the cylindrical portion to the left is continuous
with an excretory tubule. The whole cell is about 1/100th
of a millimeter in diameter. As the cilia in the cylindrical
portion undulate or beat, water is moved through the
perforations into the cavity around the cilia, and thence
into the tubule and out of the animal. Wastes are carried
along with the water (Modified from Kozloff, 1990).
|
These structures have several different designs, but basically
they are blind-ending tubes with one or more ciliated or flagellated
cells located at, or near, the blind end. The flagella or
cilia beat inside the tube's opening. In a living animal,
if viewed through a microscope, the beating of the cilia looks
rather like the flickering of a candle flame, hence the name
"flame bulb." The tube from the flame bulb connects
eventually to the animal's exterior. Although the simplest
flame bulb systems appear to be comprised of only one or two
cells, in their most advanced forms, as seen in some large
marine worms, the tubules may be quite long, comprised of
thousands of cells and many dozens, to hundreds, of flame
bulbs. The flame bulb cells are perforated near their base,
allowing water to flow through them, and the flickering of
the cilia causes water to move through the tubule to the pore's
opening to the animal's exterior.
Generally, the interior contents of the animals possessing
these protonephridia, a name meaning "first-kidneys,"
are more concentrated with dissolved materials than is their
surrounding medium. This means that water flows into the animals
through the process of osmosis. It often has been proposed
that protonephridia and, indeed, other similar so-called "excretory
organs," developed primarily to regulate animals' water
balance. These organs may indeed be primarily osmoregulatory,
but even so, much research has shown that they also function
in a manner similar to vertebrate kidneys, albeit on a very
small scale. Interior fluids arefiltered as they pass through
the perforations in the flame bulb's cells. As the filtrate
passes along the tubule, it is altered. Some materials, such
as sugars, are selectively reabsorbed, while other materials,
such as nitrogenous wastes, are actively secreted. What passes
out of the excretory pore can be called a "urine,"
quite dilute compared to vertebrate urine, but a urine nonetheless.
|
Figure 5. The protonephridia, or flame
bulbs, are often connected in complex networks, but
these are often visible only with special microscopic
techniques. At left above is a diagram of a freshwater
planarian showing its gut and the protonephridial network;
at right is a photograph, at the same scale, showing
a marine planarian. Note that its gut is quite visible,
but its protonephridia are not. Left image modified
from Kozloff, 1990.
|
|
|
|
Figure 6. A small portion of the flame
bulb network in a nemertean, modified from Kozloff,
1990. Nemerteans are among the largest animals with
protonephridial networks, and these networks are very
complex. They surround blood vessels, and the ducts
adjoining them connect and dump into larger "kidney
ducts" which have pores to the animal's exterior.
These animals may have many thousands of flame bulbs
imbedded in the tissues around their circulatory system.
|
Larger Size Brings Larger Problems
Protonephridial kidneys appear to
work fine for many smaller animals, but larger animals seem
to have replaced them with a different type of organ, called
a metanephridium or funneled organ. As with protonephridia,
metanephridia have a tubule connecting the interior business
end with "The Great Out-of-Doors." Instead of a
cell or small group of cells that create a flame bulb in the
blind-ending tube, the metanephridium ends in an open funnel
with a ciliated outer rim, called a "nephrostome,"
a term that means "kidney-mouth." The microscopic
hair-like cilia surrounding the rim beat, thereby forcing
fluid and particulate matter into the tubule. This tubule
also has areas of active secretion and resorption so that
the product passing through the excretory pore is a true urine.
Metanephridia may also have a storage bladder. One thing that
a metanephridium can do that protonephridia cannot is to sweep
small particulate materials, such as bacteria, into their
funnel, and thence eventually out of their body. It is, in
fact, an all-purpose excretory organ, removing both solid
and dissolved wastes.
Metanephridia are found in most of the larger, more complex
animals. They reach their largest size in clams such as Tridacna,
where they are called either kidneys or nephridia, but they
are nonetheless modified metanephridia. Organs structurally
identical to metanephridia but having a different name, fallopian
tubules, are found in the mammalian reproductive system. Even
the specialized and highly derived excretory organs of the
cephalopods are thought to have been derived from metanephridia.
|
Figure 7. A diagram of a metanephridium of an
earthworm (modified from Kozloff, 1990). Although the
description of the metanephridium is relatively simple,
the physiological reactions occurring in it are anything
but simple. The various loops are sites of secretion
and absorption. The kidney pore opens to the animal's
exterior, and the worm has a pair of metanephridia in
most segments.
|
Bugs
Arthropods do things much differently.
Beating cilia are lacking throughout the arthropods, probably
as a function of both their solid exterior integument and
their relatively high internal fluid pressure. As a consequence,
they don't have either proto- or metanephridia; all excretion
appears to be wholly glandular. The main arthropodan groups,
the insects, the crustaceans, and the chelicerates, all appear
to have different excretory organs. While I will discuss only
the crustacean system here, the insect system is much better
known and anyone who is interested in this topic as it relates
to insects should consult a reference book dealing with insect
physiology. The openings in the crustacean excretory system
vary in position between the different groups, but are often
found in the head region. Although such glands, called antennal
or coxal glands, have been known for years to be osmoregulatory,
and have been considered for a long time to be excretory,
little consistent physiological evidence supports the latter
contention. Until recently, these glandular structures have
been considered to be derived from metanephridia, and their
function related to them; however, given some of the recent
discoveries indicating that no arthropodan animals are derived
from an animal that could have possessed a metanephridium
or any of its precursors, it has become increasingly clear
that excretion in marine arthropods needs a lot of study before
any generalizations can be made.
A Wasteful Subject
Although the study of excretory systems
may seem far removed from the reef aquarium hobby, it is important
to remember that one of the primary problems in our systems
is the accumulation of dissolved nutrient materials. These
may accumulate from a great many varied sources, but probably
the major source of most of the dissolved nutrients in reef
aquaria is the nitrogenous waste produced by protein metabolism
in the reef's animals. Some points need to be made considering
this material. In a reef with a thriving community of small
organisms, the "clean-up crew," if you will, very
little added food is going to be subject to direct bacterial
decomposition. Almost all of it will be eaten by an animal.
As a consequence of that feeding, virtually all of the dissolved
nitrogenous nutrient found in the system must have passed
through an animal at least once.
From this state of affairs comes several important points.
First, aquarists can't rapidly or directly alter dissolved
nutrient levels by changing feeding regimes. The production
of nitrogenous waste is not directly related to feeding, but
rather the metabolism of the animals in the system. As long
as these animals are still alive, they will produce nitrogenous
wastes. Food reduction will result in there being less food
to eat, but protein metabolism, the major source of nitrogenous
waste will not be appreciably changed until the animals start
to die. While this may eventually occur due to lowered nutrient
levels, it will take a relatively long time. The fastest way
to lower dissolved nutrients would be water changes coupled
with significant removal of animal biomass. Second, lowering
nutrient levels will cause animals to starve. Starving animals
first metabolize energy storage chemicals such as fats and
sugars, but during this period, normal protein metabolism
will continue relatively unabated. Then they start to metabolize
their muscles and connective tissues. This may mean that after
a prolonged period of starvation, the dissolved nitrogenous
nutrient levels will actually rise. Subsequent to this, as
the animals die, of course, the nutrient levels will ultimately
drop. Consequently, the best way to control nutrients in animal-rich
systems is to increase exports of nutrient-rich water. This
is nature's way; nutrient-rich waters get flushed from the
reef. The next best way, considering that aquarists don't
have an unlimited water system to flush things away, is to
provide some way to convert the nutrient into biomass (algae
or animal) and export that, or to find some way to increase
nitrogenous nutrient utilization by bacteria, thus allowing
the excess nitrogen to leave the system as a gas.
|



)
)
)
)
