|
Introduction
The first article
addressed several topics, with the bulk of the discussion
focusing on global patterns of material flow within the biosphere.
The three elements we examined were carbon, nitrogen and phosphorus
due to their importance in ecosystems, and because of the
tendency for the growth and reproduction of living things
to be limited by access to these resources. This month we
will focus on factors critical to ecosystem function and conclude
with a discussion of some of the patterns in the oceanic ecosystem.
An Ecosystem Is…
I've used the term
ecosystem several times already, but I haven't really defined
what an ecosystem is. An ecosystem is a particular
assemblage of living organisms (called biotic factors) along
with the non-living components (called abiotic factors) present
in an environment that exert influence over each other. For
example, the kelp forests off North America's west coast can
be referred to as an ecosystem. Upwelling of deep oceanic
water results in a high availability of nitrogen and phosphorus.
These encourage phytoplanktonic growth as the base of a complex
food web that eventually results in productive fisheries.
This upwelling is controlled primarily by winds that blow
parallel to the coast. Without the wind there would be no
upwelling, without the upwelling there would be no abundance
of nutrients, and without the growth of phytoplankton from
the nutrients there would be no fisheries. In addition to
the importance of abiotic factors such as wind, nutrient concentrations
and currents, certain organisms live in this place and exert
influence over one another as well. Kelp provides habitat
for many of the fishes of economic interest. Sea urchins eat
the kelp, but sea otters eat the urchins. Without the otters
the urchins would multiply extensively and graze the kelp
to their holdfasts, destroying the habitat for the fishes.
Without the otters there would be too many urchins, and when
there are too many urchins there is no kelp, and when there
is no kelp the fisheries collapse. Both biotic and abiotic
factors shape the structure of an ecosystem. An ecosystem
needn't be large, either. A pool of water in a dead leaf on
the forest floor is rightfully considered an ecosystem. Countless
types of bacteria, fungi, flagellates and other microbes decompose
organic matter in the water. Ciliates feed on these microbes
and, in turn, are fed on by insect larvae. Temperature, pH,
incident light, rainfall and many other factors contribute
to the workings of this little living world. Additionally,
ecosystems contain a variety of food chains forming a food
web. A food chain is a graphic, linear representation
describing which organisms feed on which other organisms.
A food web is made up of many interconnecting food
chains. These are discussed further below.
For an ecosystem to form in any environment,
one fundamental requirement must be fulfilled: there must
be a source of energy that is useable by living organisms
and there must be organisms present that are able to utilize
that energy.
Productivity
In lay terms, if we say we've had
a very productive day, we mean that we have accomplished many
of our goals in a short period of time. An unproductive day
means we have accomplished very few of our goals during that
time. We use this term in everyday language to describe how
fast we are accomplishing our tasks, and it is used in much
the same way in ecology. In the plainest sense, the goal of
every living thing is simply to stay alive, grow to maturity,
and to reproduce. As such, the terms productivity, production
or productive are used in ecology to signify the rate at which
new materials are produced from their precursors by living
things, whether they are new tissues within an organism, offspring
or even carbohydrates that leak from a cell. The productivity
of an organism or population is the rate at which it turns
resources into more of itself. Productivity represents the
flux of nutrients from the environment into living organisms.
Often we may want to know how fast not only a single species
is growing and reproducing, but how fast all species
are growing and reproducing within a given area. One might
go to the far north and see the extensive boreal forests,
full of countless and massive trees. One might also go to
the equatorial region and see the extensive tropical rainforests
there, full of countless and massive trees. In this way the
two ecosystems appear rather similar. The trees in the boreal
forest, however, experience a very short growing season, perhaps
less than eight weeks. While the forests are vast and the
trees gigantic, they grow rather slowly as an annual community
average, as do the other plants in the forest. This environment
shows relatively low productivity. In the tropical rainforest
the growing season is year-round. Not only the trees, but
especially the forest's other plants, grow at incredibly fast
rates, much faster than those in the boreal forest. This environment
shows relatively high productivity.
The rate of production in an ecosystem bears great importance
on a variety of ecological questions. For instance, a grazing
animal might need to eat some amount of plant material per
day to be healthy and grow normally. In a highly productive
habitat the organism won't have to travel far to get sufficient
food. It can graze one area, move on to another, and then
return to the original area in short order since the vegetation
will have grown back in a short period of time. In a less
productive habitat the grazer will have to wait a very long
time before it can return to the original patch because that
vegetation grows much more slowly. This means each of this
species' grazers and any competitors for these resources must
all have larger ranges and travel more to acquire sufficient
food than they would if they lived in the more productive
habitat. The result is that less food is available in areas
with low productivity, and these habitats support fewer numbers
of individuals (as well as lower species diversity often times,
though not always) than habitats with relatively higher productivity.
Another example, and one that's important to humans, involves
natural food production. Sardines have very high rates of
productivity and can, at least potentially, be harvested at
high rates year after year. Most shark species, on the other
hand, have very low rates of productivity, and even just a
few years of intensive harvest can decimate a shark population
for decades or more. In fact, this is exactly what has happened
in most places today, though even the ultra-productive sardines
have been overharvested in some places, even to the point
of the collapse of the fishery.
Because the major source of energy for terrestrial and shallow,
aquatic environments is sunlight, the presence of plants,
algae and bacteria capable of capturing that energy is the
critical element that allows the formation of the ecosystems
with which we are most familiar. These organisms are the proximate
source of energy and most nutrients for every other organism
within the ecosystem. For this reason plants, algae and photosynthetic
bacteria are termed primary producers, because they
produce the organic material necessary to support the rest
of the ecosystem. Herbivores that feed directly on primary
producers are called secondary producers or, often
times, primary consumers. They are the second link
in a food chain, hence the secondary producers, but are the
first group of organisms that consume already formed organic
materials. Carnivores that eat these herbivores are usually
called simply secondary consumers or, rarely, tertiary
producers. Carnivores that eat these carnivores would
be tertiary consumers. These links in the food chain are called
trophic levels, with each being higher than the last
(first, second, third, fourth, etc.). Terrestrial food chains
rarely have more than four links, though exceptions certainly
exist. An example of such a chain might be:
grasses →
grasshoppers →
songbirds →
falcons
Aquatic ecosystems' food chains tend to be slightly longer
than those on land, with five to seven links, but are rarely
longer. An example might be:
phytoplankton →
zooplankton →
larval fish →
small fish →
large fish
Some might be wondering why food chains
tend to be limited to so few links. Why not have a "very
large fish" that eats the "large fish" above,
and then a "very, very large fish" to eat that one,
and then a "very, very, very large fish" to eat
that one? Certainly, the reason is not biological or physical
limitations on what can be consumed. Many fish can certainly
eat other fish only slightly smaller than themselves (or even
larger, as some owners of Antennaris spp. can attest!).
Why not have a 2 cm-long fish eat a 1 cm-long fish, and then
a 4 cm-long fish eat the 2 cm-long fish, and then a 6-8 cm-long
fish eat the 4 cm-long fish, and on up to the size of white
sharks? Why not have hundreds of links in a food chain? Ahh,
good question young student-of-life-as it turns out there
is a very important reason this does not happen, and it is
explained by the Second Law of Thermodynamics.
The Second Law of Thermodynamics
"The entropy of an isolated system will tend to increase
over time approaching a maximum value."
…so what the heck does that mean?
A simpler way to say this is that all natural systems tend
to move from order (everything is arranged nicely) to disorder
(everything is completely jumbled and homogenized) without
the input of energy. Think of a nice, ordered playroom where
all the toys are put away in bins, the paint is in the jars
and the room is spick-and-span. Everything has its place and
everything is in its place; there is order. What happens after
releasing some children into the room for awhile? Disorder!
The toys go everywhere, the paint inevitably ends up on the
walls, and everything becomes thoroughly disorganized. No
matter how long we wait, the toys will not go back into their
bins by themselves, the paint will not clean itself off the
walls, and the room will not become organized without an input
of energy to make all of this happen. Undoubtedly, the input
comes from some frustrated parents, but I digress. The important
point to glean from this is as follows: everything in nature
moves from order and organization to disorder and homogenization
without the input of energy to prevent it from happening.
Another consequence of this law is that
no transfer of energy is completely efficient. When wood is
burned in a campfire to roast marshmallows, some of the energy
is released as infrared radiation, which does the roasting,
but a great deal is also lost into the environment as unused
infrared radiation (i.e., infrared that does not directly
roast the marshmallow), some is lost as light, some is lost
as sound, etc. What this means is that every time the energy
is converted from one form to another, some of it is lost
to the environment (entropy is increased).
How does this help us to understand ecosystem
ecology? First, living things are ordered structures in a
sea of chaos. Almost everything about a living cell distinguishes
it from its environment. Let us consider a human body. Mostly
it is made of water, yet this water tends to quickly evaporate
into the air in which we live. Our body contains salt, yet
this salt is not leaked into the water if we swim in a lake,
nor are we flooded by salt if we swim in the salty ocean.
We maintain body temperatures at 37° C (98.6° F) even
as the temperature around us rises above, and dips well below,
that temperature. The proteins in our bodies maintain their
shapes, even as chemical and physical laws work to break them
apart. Maintaining this order, that is, being unique from
the environment that surrounds us, requires a constant supply
of, and a great deal of, energy. Much of the energy in the
food we eat, however, is lost to our environment because no
energy transformation from one form to another is ever 100%
efficient. We lose a great deal of energy as radiant heat
to our cooler environment, as sound while performing various
tasks, as mechanical energy as we move objects, etc. We also
lose some of the energy in our food as undigested material.
For example, we and most organisms can't digest cellulose-fiber-and
therefore gain no energy by eating this material, because
it is not digested, though it took the original capture of
a great deal of energy from the sun to produce that cellulose
by whatever plant made it. With every step up a food chain
there is less and less of the energy that was originally captured
transferred to the next trophic level.
So, how much energy is transferred from
one trophic level to the next? The true value depends on many
factors, but for most organisms this value is about 10% of
the energy they consume. In other words, to produce 1 lb.
of cow you need about 10 lbs. of grass. In order to produce
1 lb. of salmon you need about 10 lbs. of small fish, about
100 lbs. of zooplankton to produce the small fish and about
1000 lbs. of phytoplankton to produce the zooplankton. Only
five links require 10,000 lbs. of primary producer to produce
1 lb. of top predator, 100,000 lbs. with six links and 1,000,000
lbs. of primary producer to produce a single pound of top
predator with a seven-link food chain! Now we can appreciate
why food chains longer than this tend to be so rare. Usually,
food chains longer than seven links (which do exist, especially
in the ocean) are able to exist only because one or more of
their links are far more efficient producers than those in
the above examples, hence more energy is passed on than otherwise
would be.
Oceanic Productivity
We understand now that productivity
is a very important parameter influencing how an ecosystem
functions. The logical question for all of us is how productive
are the oceans, especially the tropical oceans where coral
reefs grow? A multitude of factors influence precisely how
to answer this question, but in general we can say, "not
very," especially as compared to many other ecosystems.
Typically when we discuss the productivity of an ecosystem
we mean its net primary productivity, that is, the sum of
all net production of all species of primary producer; in
other words, how fast the plants, algae and photosynthetic
bacteria in a given area are growing. Often this value is
expressed as the dry weight of production in grams of carbon
per square meter per year (g C m-1
yr-1), though it might also
be expressed in some other set of units (e.g., grams of carbon
per square meter per day) depending on the information being
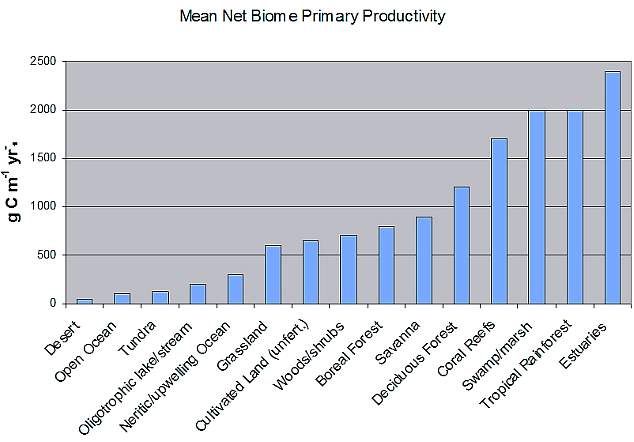
presented. Figure 1 shows the mean rates of primary production
for a number of biomes. These are average values and some
geographic variation is to be expected.
Notice that the open ocean is at the far left. It is one
of the least productive of the world's major biomes over a
given area (though it is by far the largest biome and therefore
contributes a major portion to worldwide productivity, despite
its own poor rate of productivity, because of its immense
size). Why is the open ocean so relatively unproductive? Actually,
the reasons are rather simple; so simple, in fact, that two
boxes with a few lines drawn between them has proven to be
a good model of what is happening in this complex system.
|
Figure
1. Mean net primary production in selected ecosystems.
|
A Bit of Oceanography
If a person swims
in a lake in the temperate zone during the summer and dives
into deeper water, he or she will probably notice that the
surface water is very warm and abruptly becomes several degrees
cooler as he or she descends, and then remains at this cooler
temperature. In fact, during the summer this lake will become
stratified such that a thin layer of warm water is floating
on a thick layer of cold water. While water is obviously quite
fluid and it seems as though the two layers should mix easily,
just the opposite happens. A thin zone called a thermocline
is established where the temperature changes abruptly. It
is almost as if there are two separate water bodies, one over
the other. Things that swim, sink or float can move between
these two layers easily enough, but anything that moves with
the water (such as dissolved substances like the nutrients
necessary to fuel algal growth) can and does become trapped
in whichever layer it happens to be in before the thermocline
is established. During the Fall the top layer is not heated
as much by the sun as it is during the summer, so the thermocline
begins to break down and the water layers begin to mix. During
the Winter there is no thermocline at all and the water becomes
well mixed. During the Spring the thermocline begins to reestablish
and the lake again begins to divide into two separate layers.
This same general pattern is also observed in the ocean, though
its large size allows this phenomenon to manifest in at least
three different ways.
In the polar oceans the sun is strong enough
to establish only a slight thermocline during the summer and
no thermocline during the rest of the year, so the surface
and deep water tend to stay well-mixed throughout the year
and not to stratify. The temperate seas tend to follow seasonal
stratification and mixing scenarios like those described above.
In the tropics there tends to be a constant thermocline due
to the perpetual heating by the sun, and thus the surface
and deep waters tend to be constantly separated. This thermocline
occurs rather deeply in the tropics, at approximately 100-200
m (330-660 ft.), in other words, deeper than any of the animals
we might normally keep in a reef aquarium. I must point out
that this oceanic thermocline is distinct from the transient
thermoclines that sometimes become established in shallow
water and might be felt by a diver as they descend on a reef.
The two are very different. The oceanic thermocline could
be approached only by a technical diver on a very deep and
potentially dangerous dive. Additionally, there is relatively
slow mixing between polar and temperate oceans and temperate
and tropical ones on the surface. Because coral reefs are
found only in shallow, tropical waters, the ocean around coral
reefs can be treated as a semi-isolated water body with slow
rates of mixing with the rest of the ocean, though certainly
mixing does still occur.
Again, particles can move between these
layers if they sink, float or can swim, but dissolved substances
are essentially trapped. One of the hallmarks of the open
ocean (often called the oligotrophic ocean) is the
extreme scarcity of dissolved nitrogen and phosphorus sources
that can be used by primary producers. Furthermore, the standing
stock of primary producers is extremely low. Higher
trophic levels are only very slightly more abundant than lower
levels in terms of standing stock. The shallow, tropical ocean
is devoid of neither life nor nutrients but, compared to many
ecosystems (especially terrestrial ecosystems), it has a relatively
low concentration of both inorganic and organic nutrients.
On the contrary, deep oceanic water anywhere in the world,
including in the tropics, tends to be quite rich in inorganic
nitrogen and phosphorus that can be used by algae (though
there is insufficient light at these depths for algae to grow).
We might ask why there is a large supply of nutrients in the
deep water and a short supply in the shallow water. It is
for two reasons: 1) the lack of mixing between deep and shallow
water as explained above, and 2) the biological pump.
The Biological Pump
When a phytoplankter
takes up inorganic carbon, nitrogen and phosphorus from its
environment (and any other nutrients or materials, for that
matter), it reduces the pool of these dissolved nutrients
in the water that surrounds it. These nutrients are used for
the growth and maintenance of that algal cell and when the
alga acquires enough nutrients, it splits into two daughter
cells. Both of these daughter cells take up nutrients from
their environment, further lowering the available supply in
the water. They then grow and reproduce, splitting into four
cells. Then these four cells continue their activities just
as their parent cells did. Given the growth rate of most phytoplankton,
it is easy to see that they will quickly draw down the inorganic
nutrient supply in the water and fill the water instead with
algal cells. While they are alive these phytoplankton tend
to remain neutrally buoyant through various flotation mechanisms,
but many are negatively buoyant without the expenditure of
energy. Thus, when they die they begin to sink. The same is
true of most other forms of life such as zooplankton and nekton
such as fish and whales. Most of this production is decomposed
in the water column and mineralized into inorganic components.
How quickly a particle is decomposed (primarily by heterotrophic
bacteria) depends primarily on its size and how labile
it is. A tiny algal cell will be decomposed much more quickly
than a Humpback whale, for example.
If either a phytoplankter or a whale
die at the surface, they will begin to sink and bacteria will
begin to decompose them. As they are decomposed the organic
molecules they are made of will eventually be reduced to inorganic
ions (nutrients) and released into the water, where they can
be absorbed and reused to fuel the production of more phytoplankton.
The algal cell will likely be completely or almost completely
decomposed before it reaches the thermocline. There will have
been a flux of nutrients from the water to the alga as it
was born and grew, and then back to the water after it dies
and decomposes, but there will be no net addition or loss
of any nutrients from the surface waters. The whale, on the
other hand, likely will not be totally decomposed before it
reaches the thermocline, but will keep on sinking and decomposing
until it is totally decomposed in the water column, or reaches
the bottom and is fully decomposed. Either way, it will carry
a great deal of nutrients from the surface below the thermocline
in the form of a particle. The same happens if the phytoplankter
dies nearer the thermocline-it sinks to the deep water layer,
and takes all of its sequestered nutrients with it, before
it is fully decomposed. These nutrients are mineralized into
dissolved substances which do not freely move across the thermocline.
In this way the surface waters lose nutrients to the deep
ocean. It should also be noted that this allows carbon to
be lost from the surface ocean and sequestered in the deep
ocean. This mechanism is important to the global carbon cycle
and future climate projections. An important point to stress,
and one that is often misunderstood, is that particles do
not have to reach the bottom of the ocean to remove nutrients
from the surface layer; they simply have to sink below the
thermocline. The shallow, tropical ocean is, therefore, constantly
losing nutrients to the deep sea if these particles are able
to sink below the thermocline. Because these layers mix very
little, the dissolved, inorganic nutrient pool in shallow
water is not recharged seasonally as it is in the temperate
zone, or constantly as it is in the polar seas. Therefore,
the tropical ocean tends to have perpetually low concentrations
of dissolved inorganic nitrogen (DIN) and dissolved inorganic
phosphorus (DIP), unless local conditions allow a temporary
influx of these nutrients from land, the atmosphere or deep
oceanic water (these sources will be discussed in upcoming
segments). The productivity of the oligotrophic ocean, especially
in the tropics, is therefore low due to nutrient limitation
on the phytoplankton. But which nutrient is limiting them,
and why?
Redfield's Ratio
Alfred C. Redfield
was teaching physiology at Harvard Medical School when he
was recruited by Henry Bigalow as a staffer for the newly
formed Woods Hole Oceanographic Institute. He split his time,
teaching at Harvard during the academic year and doing research
at Woods Hole, for more than ten years, until 1942 when he
took a one year leave of absence from Harvard (which actually
lasted 14 years) to become the Associate Director at Woods
Hole.
It was during his work at Woods Hole that he made an observation.
The molar ratio of N:P in phytoplankton tends to be very similar
to the molar ratio of DIN:DIP in deep oceanic water. He hypothesized,
correctly, that it is not the availability of these nutrients
that determines the make-up of phytoplankton, but rather the
make-up of phytoplankton that creates this ratio in the water.
The molar ratio he found for carbon, nitrogen and phosphorus
is 106:16:1-106 atoms of carbon for 16 atoms of nitrogen for
1 atom of phosphorus in an average sampling of phytoplankters
(Redfield, 1958). This is called a stoichiometric relationship.
You may recall the word "stoichiometry" from your
high school chemistry class (or perhaps not, depending on
how well you have blocked out those memories!). In chemistry,
stoichiometry is often introduced when students are taught
to balance chemical equations. Stoichiometry refers
to the relative proportion of constituents in a material.
Likewise, the stoichiometric relationship between car wheels
and drivers is 4:1 whereas that of motorcycle wheels to drivers
is just 2:1. Understanding the stoichiometry of living organisms
and how this impacts their interactions with each other and
with their environment is often referred to as ecological
stoichiometry. To be clear, these are molar ratios, not
mass ratios. The mass of a phosphorous atom is more than double
the mass of a nitrogen atom. Thus, a molar ratio of 16:1 N:P
becomes nearer 7:1 as a mass ratio. This is an important point
to consider, as hobbyists usually measure dissolved nutrient
concentrations in parts per million (ppm), which refers to
mass, not moles.
Misunderstanding Redfield's Ratio
Unfortunately, all
too often Redfield's ratio of 106:16:1 atoms of carbon, nitrogen
and phosphorus has been interpreted not as the outcome of
the interaction of thousands of species of phytoplankters
over many generations' worth of time, but rather as the stoichiometric
relationship to be expected in all phytoplankton (or worse
yet, all algae) and all oceanic waters all of the time. Simply
put, this is a gross misinterpretation. Some species of phytoplankton
normally have N:P ratios of more than 45:1 (Klausmeier et
al, 2004). Others might have N:P ratios as low as 8:1
(Klausmeier et al, 2004). It is the aggregate response
of entire communities over many generations that produces
the observed pattern. In fact, Atkinson and Smith (1983) found
that benthic
algae like those that grow on coral reefs or in our tanks
tend to have a mean ratio of 550:30:1 atoms of C:N:P. Therefore,
we should not expect the water in our tanks to always have
molar N:P concentrations of 16:1, or even 30:1, or that variance
from these ratios necessarily represents either nitrogen or
phosphorus limitation of the algae in our systems. Not only
do different species of algae grow with different N:P ratios,
so do the same species, within a range of tolerance. I have
seen hobbyists with slightly elevated DIN levels and barely
detectable DIP levels dump significant amounts of various
phosphate supplements into their tanks in an attempt to alleviate
P-limitation and cause the algae to consume the nitrogen.
They didn't drop their DIN concentrations, but they did cause
significant damage to their corals and their tanks in general.
There is no reason to think that our tanks should precisely
reflect these ratios all of the time. In fact, N:P ratios
of 10:1-20:1 for phytoplankton are often considered to represent
cases of co-limitation, because most phytoplankton that can
grow with a 16:1 ratio can easily grow in the range of 10:1-20:1
(Hamilton, pers. comm.). Addition of either N or P often stimulates
growth of phytoplankton in such a situation…except in
the Southern Ocean.
The Southern Ocean is both temperate and
subpolar in parts. It contains plenty of available nitrogen
and phosphorus to stimulate significant phytoplanktonic growth
during the warmer months of the year, just like in the Northern
Hemisphere, yet this is the least productive part of the ocean.
As I mentioned last month, carbon, nitrogen and phosphorus
are most frequently the limiting nutrients for organismal
growth, but they're not the only nutrients that can
limit growth. Nutrient limitation occurs when the supply of
any required nutrient is low relative to how much is
needed. If we extend Redfield's ratio to include iron, we
find that phytoplankton tend to have a stoichiometric relationship
of 106:16:1:0.005 atoms of C:N:P:Fe. The primary productivity
of much of the Southern Ocean is limited by the availability
of iron (Behrenfield and Falkowski, 1999; De Baar et al,
1990; Martin et al, 1991).
If nothing else, this must remind us as
reef aquarists that not every interaction involving algae
or nutrients in our tanks is simple or necessarily obvious,
and that nitrogen and phosphorus, while very important pieces
of the puzzle, are still just two pieces of a large and complex
whole. Idiosyncratic interactions in ecosystems as diverse
as a coral reef are commonplace, and if we are to appreciate
these interactions and how we can better maintain our tanks,
we must consider them.
Where do we go next?
My goal in the rest
of this series is to bring to light not only the simple and
well established interactions between nutrients, algae and
higher organisms on a coral reef, but also to draw out many
of the complex or little-known influences that each has on
the other, and to make this information accessible to average
reef hobbyists in ways that will improve how they design and
maintain their own reef aquariums on a day-to-day basis.
Some may have noticed in Figure 1 that,
while the mean net primary productivity in the oligotrophic
ocean surrounding coral reefs is fairly low, the mean net
primary productivity of a coral reef is much, much higher-an
order of magnitude higher, in fact! How can that be? Why are
coral reefs so productive over a given area, when the ocean
around them is so unproductive? How do they maintain such
large and complex food webs? Next month we will begin to answer
these questions and to better understand how and why our aquariums
function.
|