|
"If all the matter in the universe except
the nematodes were swept away, our world would still be dimly
recognizable... we would find its mountains, hills, valleys,
rivers, lakes and oceans represented by a film of nematodes."
Nathan Augustus Cobb, 1914
Worms' Worms
Given the immense
variety of animals, it might appear difficult to pick a single
type of animal as being characteristic of all the Earth's
animals. The diversity of animal life is not, however, distributed
equally across all the various "types" of animals,
and some types include many more species than others. These
species-rich animal groups surely are more "characteristic"
of all animals than are those groups containing only a few
different varieties. Each distinctive "type" of
animal life may be considered to be the taxonomic groups called
"phyla" (singular = phylum). A phylum is an arbitrary
grouping of similar animals thought to represent a unique
and specific "body plan" or architecture. Most taxonomists
consider between thirty and fifty named phyla as constituting
the complete array of animals' body plans. These represent
the gamut of animals' structural patterns, from the simplicity
seen in a sponge or coral polyp to the profound structural
complexity seen in a blue whale or a butterfly's caterpillar.
In some ways it is easy to array all of these various body
plans in some sort of order, from simplest to most complex,
for example, and compare them. Because such comparisons are
so easy, this process has been hammered to death by zoologists
for the last couple of centuries, with the objective of trying
to find some sort of order in life's immense diversity. Visually
examining and comparing the body plans results in… chaos,
confusion, argument and resignation. There is no clear way
to simply look at, examine and arrange the various phyla in
an inherently obvious and rational arrangement. Instead, there
are a LOT of "inherently obvious and rational arrangements,"
some of which are more "inherently obvious" than
others. And, all of these various arrangements vary between
being somewhat different to radically different from each
other, and all of these constructed and artificial patterns
depend upon assumptions made during the comparisons and orderings.
Assumptions of this nature are always wonderful things, as
they are based on those delicious and delicate ambiguities
known in the vulgarity of college life as "weasel words,"
and in the morass of national politics as "opportunity."
As an example, while it is easy to conceptualize the concept
of "complexity" with regard to animals, it is well
nigh impossible to erect some precise, unambiguous and universally
agreed upon "measure" or standard of complexity.
Visually comparing two very different, yet complex, body plans,
such as the one for arthropods and the one for fishes, is
very difficult, as they share almost no structural similarities.
Both designs lead to very complicated animals, but in very
different ways. There is really no way to compare such "finned"
apples and "clawed" oranges to come up with a relative
index of complexity. Problems such as this caused stagnation
of research into the relationships between the various animal
groups; from about 1940 until the late 1990's few new ideas,
and almost no new evidence, were presented about how animal
groups were interrelated. There was a lot of passionate discussion
about their relationships, generally accompanied by some serious
waving of arms, but as nobody had any unambiguous data, independent
of conjecture, there was no way to resolve the various competing
hypotheses. All of this changed with the development of genetic
comparisons, whereby various components of the actual genetic
material that determined the structures could be compared.
This allowed for the determination of which animal groups
had the most similar genetic material or, in other words,
which groups were most closely related. Applied within a wide
series of group-by-group comparisons, this process has facilitated
the development of a
framework of relationships that is based on both genetic
and structural characteristics relating most major animal
groups to one another. Nonetheless, even with this group of
methodologies, there are still some unanswered questions,
both of technique and result.
Such a framework of relationships shows which animal types
share lines of descent from common ancestors. Applying some
biological acumen to the changes that occur when lineages
diverge can, however, also lead to an appreciation of the
ways in which animals have come to exploit different environments.
Generally, the more complicated the animal's basic body plan,
the more different types of habitats in which the animals
sharing that plan can exist. It is generally conceded that
arthropods, chordates and mollusks share a level of complexity
not reached by other animals. Consequently, and not surprisingly,
arthropods, mollusks and chordates are found in just about
every habitat on Earth. Animals with a simpler level of organization,
such as sponges or corals, are much more restricted in the
habitats they can occupy. As a result, we might suppose that
the animal considered to be most characteristic of all animal
life would be either an arthropod, a mollusk or a chordate.
Perhaps, but there is another option.
The worms in the Phylum Nematoda constitute the one notable
exception to this relationship between structural complexity
and diversity of habitats occupied. These worms, commonly
called "round worms," are fundamentally simple in
structure and, yet, are not only found almost everywhere,
but also are found in just about every other type of multicellular
living organism. It is quite possible that there are more
different species of nematodes than of all other animals combined.
Because of their widespread existence, it might reasonably
be said that the most characteristic and ubiquitous form of
animal life is a nematode. The nematodes' body plan is truly
the design of a worm's worm.
|
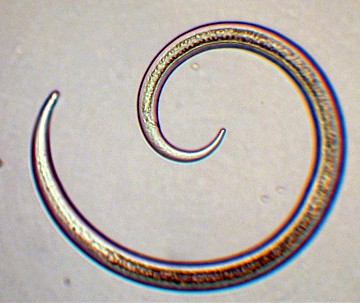
Figure 1. An unidentified free-living nematode
found in marine sediments. Its basic structural simplicity
is evident.
|
Why?
Given that nematodes live everywhere
and in everything, it is reasonable to ask, "Why?"
What special properties or characteristics that nematodes
alone possess have allowed this degree of diversity? When
a group of organisms has diversified into many different forms,
this is referred to as an "adaptive radiation."
The organisms' group appears to have spread from one central
area, or diversified from one ancestral type, much as light
radiates from a single source. In the process, as they have
encountered new habitats or situations, the organisms have
adapted to them.
|
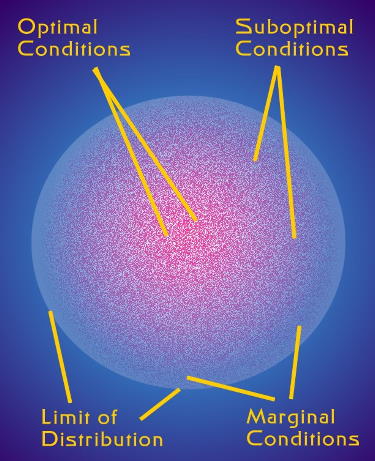
Figure 2. The process of adaptive radiation needs
populations of organisms spread over a wide variety
of conditions. New species typically arise when organisms
living under suboptimal or marginal conditions undergo
genetic changes that allow them to exploit those conditions
more fully than their ancestors.
|
Diversification of animals is not an automatic process, and
a good many groups have never diversified very much. Typically,
diversification requires some properties of form and function
that allow exploitation of new habitats or life-styles. One
of the reasons that there are so many species of some types
of animals such as insects, snails or fishes is that their
body's form is "plastic" or "malleable"
when subjected to evolutionarily selective pressures, and
it can change or form into new shapes that are adapted to
particular habitats or lifestyles. Such "variable"
organism groups may also have more genetic material that may
be modifiable, which in the end, may result in structural
variations.
That type of diversification appears to be exactly the opposite
of what has occurred in the nematodes. The great diversity
seen in the nematodes notwithstanding, the nematodes are a
VERY homogeneous group with regard to morphology. Unlike
many of the more complicated types of animals, the key to
nematodes' diversity appears, in many ways, to be their structural
simplicity, coupled with a design that allows the exploitation
of an extremely common environment. The nematode body "design"
appears to have been "preadapted" to the exploitation
of many environments.
The Worm's Outside Surface
The nematode's body is a thin cylinder
which tapers to a fine point at either end. The common name
"round worm" refers to these worms' cross-sectional
appearance, which stands in contrast to the many "flatworms"
seen so commonly in marine aquaria and elsewhere. Their lack
of any indication of true segmentation separates and distinguishes
them, as well, from the annelid worms, such as "bristle"
and earth worms.
The outermost covering of a nematode's body is a non-living
proteinaceous cuticle, and although some species have a superficially
annulated cuticle, they lack the internal divisions characteristic
of truly segmented animals. Nematodes also lack any appendages,
although they may possess sensory bristles or hairs. The nematodes'
cuticle is a complex and multilayered structure that is secreted
by an underlying epidermis. The cuticle's major layers consist
of the protein collagen, which also is the major constituent
of vertebrate ligaments. Collagen is NOT elastic and NOT stretchable.
Generally, at least three layers of collagen are found secreted
tightly adjacent to one another in an arrangement not unlike
the layers of wood in plywood or some other laminate. The
"grain"
of the various layers alternates in direction, giving
strength and resiliency to the body wall.
Visible external structures are relatively few. Nematodes
have a few complex sensory organs, but eyes are rare. Occasionally,
when eyes are present, they are found inside the throat. Other
sensory structures are located on various bumps and pits in
the cuticular surface. Most of these structures are innervated
and are presumed to be sensory, but the exact stimuli that
they respond to are unknown. Most researchers presume that
they possess both tactile and chemosensory capabilities. Nematodes
may also possess "sensory" pits called "amphids"
at either side of their body just at the back of their head,
but their sensory modality is unknown. Depending on the species,
there may be additional papillae in the "cervical region."
Additionally, some nematodes have structures called "phasmids"
which appear to be adhesive organs. Phasmids may be duo-gland
adhesive structures. As the name "duo-gland" implies,
these structures consist of two glands working together. When
the animal wants to remain attached to something, one gland
secretes an adhesive, and when it wants to move, the other
secretes a releaser substance that dissolves the adhesive
and the worm will be on its way.
It is worth noting that some of these descriptive terms are
rather ludicrous. Nematodes have neither a defined head nor
a neck; however, both terms are still applied to them. Most
free-living nematodes are transparent, with internal divisions
that do delineate something that could be called an anterior
region containing the mouth's parts; that part of the animal
is referred to as the head. The area just behind this head
is, of course, the neck or cervical region.
The arrangements of exterior structures such as the papillae,
and the presence or absence of amphids and phasmids, constitute
some of the major taxonomic characters used to discriminate
the phylum's various subdivisions. To identify species or
other taxonomic subdivisions requires examination of these
structures, and they may be VERY hard to see. Most folks just
give up at this stage if they are trying to identify the worms,
and just call 'em all "nematodes."
|
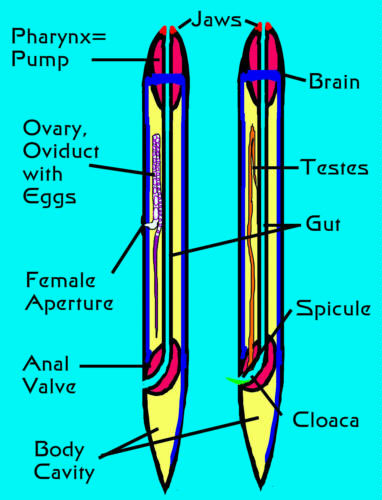
Figure 3. This is a diagrammatic representation
of the anatomy of a typical nematode; left: female,
right: male.
|
|
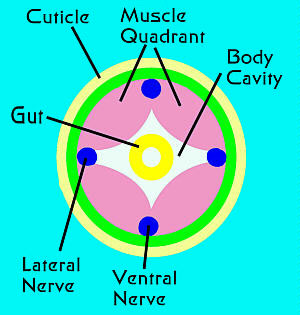
Figure 4. Diagrammatic representation of the
cross-section of a typical nematode, showing the relationships
of the body's major structures, with the exception of
the gonads.
|
Muscles and a Liquid Skeleton
Underlying the cuticle is the epidermis,
sometimes called the "hypodermis," a layer that
marks the animals' actual living surface. Inside the epidermis,
most of the remaining body wall structure is composed of muscles.
No muscles run around the body of any nematode; their only
muscles run the length of the worm. Much of the worm's structure
is dictated by this musculature. Any muscle can only actively
contract. Once contracted, all muscles must be actively stretched
back out to their expanded length to be able to contract again.
In animals such as ourselves or crabs, this expansion is done
by that system of levers we call our skeletons. In animals
with a lever-action skeleton, each muscle is paired; for each
muscle that contracts, another muscle must, by its contraction,
force the lever to pull the first muscle back to full extension.
A good example of muscle pairing is the biceps and triceps
pairing in humans' upper arm. The biceps contract to flex
the elbow, and stretch the triceps, and vice versa.
To stretch their muscles back to full extension, nematodes
do not use levers, but instead use water enclosed in a bag
and held under high pressure. Such an enclosed water volume
used to antagonize muscles is called a "hydrostatic skeleton,"
and nematodes probably have the most well-developed, simple
hydrostatic skeleton in the animal kingdom. The fluid is enclosed
in the animals' body cavities and contained there by the strength
of the body wall. This is an extremely high pressure system;
in large worms, the internal pressure can exceed 250 mm Hg
(more than normal human blood pressure).
The hydrostatic skeleton is maintained by two major factors.
First, the complex and non-expandable, non-elastic cuticle
acts to contain the body and keep it from expanding. The second
factor that is necessary for this system's functionality is
the physical process of osmosis. Osmosis is simply the diffusion
of water across a membrane in response to a water concentration
gradient. Because the worm is filled with fluid comprised
mostly of water, but also containing various chemicals, water
is less concentrated inside the cell than outside it. Hence,
water tends to ooze into the worm's body cavity. The collagen
layer and osmotic pressure tend to provide pressure to "fully
inflate." Fluid enters the worm by osmosis, and is held
back only by the physical properties of the cuticle. If the
cuticle were not present, and the worms were otherwise the
same as normal, they would explode from the influx of water.
The muscles in nematodes are arranged into four groups, one
for each quadrant of the animal. If the nematode is visualized
in cross-section, it will be seen that right and left dorsal
and ventral muscle groups run the length of its body. These
dorsal and ventral muscle groups are enervated separately
and are contracted separately in an alternating sequence to
produce the characteristic dorso-ventral flexion that constitutes
the ONLY motion available with the pattern of muscles found
in these animals. This whip-like up-and-down motion, with
the animal flexing first one way and then the other, is characteristic
of nematodes. Nematodes can't twist and turn because they
have only longitudinal muscles, and they can't move by alternately
contracting and expanding various sections of their body's
musculature. They can perform only an all-or-nothing dorsal
or ventral flexion.
This system is functionally very simple. When a muscle group
contracts, it tends to shorten the body on the top or the
bottom. This, in turn, tends to reduce the body's volume,
but as the fluid filling the body is incompressible, the volume
must remain the same. Consequently, internal pressure increases,
the opposite muscle band extends, and the worm bends. The
antagonists of the contracted muscles are the muscles on the
animal's opposite half, and the force is transmitted by the
hydrostatic skeleton maintained and coupled with the cuticular
fibrillar arrangement.
To reiterate, the only motion that nematodes can do is to
flex, either in the up (dorsal) or down (ventral) direction.
They simply cannot move in any other manner. Such motion is
inefficient for movement unless the animal is in a viscous
medium (such as a sand bed, soil, tissue
or bodily fluids) or fastened to substrate. In such media,
however, this motion is quite efficient, and people who have
some large nematodes (Ascaris) living
in their guts often can feel them actively swimming from
place to place. The efficiency of such movement in viscous
media is also the answer to the puzzle of why there are there
so many kinds of nematodes. The answer is simply that these
animals are superbly designed to move in viscous substrates,
and these substrates are everywhere, from all aspects of the
soft-sediment marine environment to terrestrial soils to the
tissues found in all animals and plants.
Nerves
As befits an animal with such a simple
set of locomotory motions, the nervous system is simple. It
consists of a nerve ring around the foregut with a ventral
swelling called the brain. Nematodes also have four nerves
running the length of their body: a dorsal nerve, a ventral
nerve and a lateral nerve at the midpoint on each side. If
you examine a worm cut perpendicularly to its long axis (termed
a cross-section), you would see that the nerves divide the
animal into equal quadrants. The lateral nerves are largely
sensory, and the dorsal and ventral nerves are largely motor
nerves conducting impulses that cause muscular contraction.
So, the lateral nerves conduct impulses to the brain, while
the dorsal and ventral nerves conduct impulses from the brain
to the muscles. Located ventrally on the anterior nerve ring,
the brain is a swelling of nerve tissue referred to as a ganglion.
It is doubled at its anterior end. There is an additional
aggregation of nerves, or ganglion, at the posterior end called
the anal ganglion.
Simple Innards
Nematodes have a simple gut. Their
mouth often has biting jaws, and in parasitic forms they may
actively bite their way through tissue. The free-living forms
use their jaws to catch prey or to grasp sediment that is
eaten. The remainder of the gut is a simple tube, only one
cellular layer thick, without folds, loops, pouches or membranes
connecting it to the body wall. It runs from the mouth-throat
region at the animal's anterior end to the rectum at its posterior
end.
The high hydrostatic pressure presents problems for the animal;
simply put, this pressure tends to force contents out of the
gut. Consequently, a rather complex system of valves controls
nutrient passage. Food has to be actively pumped into the
gut, and the throat region, called the pharynx, is basically
a muscular pump designed to do just that. Defecation occurs
by the relaxation of some of the musculature near the anus.
High internal pressure then expels the feces. Due to the high
internal hydrostatic pressure in large worms, defecation can
be an impressive event; a large pig or human round worm of
the genus Ascaris can spray its feces about 10 feet
vertically into the air or about 30 feet laterally! These
animals normally live in the host's small intestine, and in
these situations, the feces travel only a short distance from
the animal. Other than the muscular valves around either end,
the gut is simple, consisting of just a single digestive epithelial
layer, with no muscles lining it. Both extra- and intra-cellular
digestion occur, and nutrients travel from the gut to the
body wall in the fluid of the body cavity. Nematodes secrete
ammonia as their primary waste product. Most nitrogenous wastes
exit through their gut.
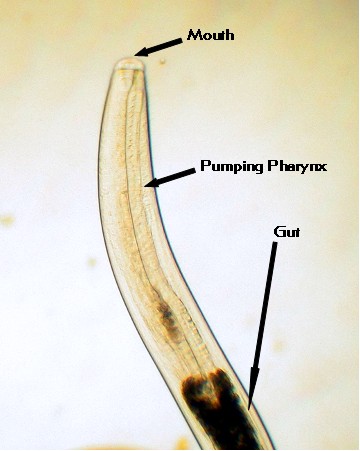 |
|
Figure 5. The anterior end of a small marine
nematode showing the muscular pharynx (throat region)
necessary to pump food into the gut.
|
Reproduction and Development
The sexes of nematodes are separate
and visually distinguishable. The male worms have a cloaca,
a common opening for the anus and the male genital pore. This
is located slightly in front of the worm's posterior end.
The male generally has a pair of copulatory spicules which
protrude from his cloaca. These assist in holding the mate's
genital aperture open during copulation. The female genital
aperture is located about midway along the animal, on her
ventral surface. As far as I know there are no hermaphroditic
nematodes. All fertilization is internal, and the male often
has accessory structures for pumping the sperm in against
the female's high internal pressure. Cilia don't work under
high pressures, and the sperm are amoeboid.
The embryological development is extremely determinate; this
means each cell's fate is determined prior to fertilization.
After each cell division, the chromosomes undergo diminution
- they get broken and reduced except in germ cell lineage.
Once development is complete, cell division ceases, except
in the gonads. No bodily, or somatic, cell can be stimulated
to divide again, and this means the animal has no way of repairing
injury or damage. Nematodes are "eutelic;" this
means that EVERY individual of a given species has EXACTLY
the same number of cell nuclei in EXACTLY the same position
in its body. For example, all individuals of the nematode
version of a "lab rat," Caenorhabditis
elegans, have 959
somatic nuclei. Interestingly enough, while they may have
"cell" nuclei, what they lack are actual cells.
Most bodily structures in nematodes are syncytial; the nuclei
that normally would be expected to exist within cells are
found in masses of protoplasm, but no cell walls delineate
actual cells within these masses. Lacking cells and cellular
division, nematodes are incapable of repairing injury and
if damaged, they will die.
As in the arthropods, the cuticle constrains growth, so to
increase in size, the animals must molt. Most nematode species
have five life history stages (resulting from four molts).
Growth is achieved primarily by an increase in cellular size,
and the animal's size is NOT proportional to molt stages.
Many species remain very small until their last molt and then
increase significantly in size. Round worms are taxonomically
placed into the Phylum
Nematoda, in the branch of the animal kingdom called the Ecdysozoa,
as they must molt to grow. The term Ecdysozoa is derived from
roots meaning animals that "strip" or molt, hence
all animals that molt are put into this group. While Cobb
in his quote above was undoubtedly not thinking of marine
reef aquaria, the quote that started this article is just
as undoubtedly applicable to them. Nobody really has an honest
clue as to the absolute diversity of nematodes; their basic
structural similarity makes distinguishing species an absolute
nightmare. Nonetheless, what appear to be reasonable estimates
of the number of species range upward to over 10,000,000.
An Identification Nightmare
The apparent consistency of form exhibited
in the group is so overwhelming that nonspecialists often
say that "There is only one nematode species; it just
comes in different sizes." This
is not at all true, (2),
but the differences between the species often appear to be
exceptionally subtle, and it takes thorough training to be
able to distinguish many species. Some of this may be due
to our problems in defining species by the appropriate criteria.
Although almost all biologists give lip service to the "biological"
species description of Ernst Mayr which states that "a
species is an interbreeding unit," in practice, most
biologists fall back onto the "morphological" species
description.
Basically, the "morphospecies" concept implies
that if there are consistent and significant differences between
individuals from two populations, then these two populations
are two species. Or... phrased another way, "If I can
tell 'em apart, they come from two species; if I can't tell
'em apart, then they form one species." There are some
obvious problems with this approach, not the least of which
is that it totally ignores the animals' actual biology. Additionally,
it depends upon a subjective "feeling" of difference.
In other words, how far apart do two animals have to be to
be in different species, and just exactly how do taxonomists
measure that magnitude of difference?
Keeping these little problems in mind, you have to realize
that we know very little about the natural history of any
free-living nematode, and that coral reef dwelling forms are
particularly poorly understood and very difficult to tell
apart. Because of this we have to fall back on the morphological
species concept. We can't use the biological species concept
simply because we don't know enough about their biology. Nematologists,
fortunately, seem to be a rather conservative crowd, and seem
to require a lot of differences between their animals before
they refer to them as two different species. That magnitude
of differences should help us identify the beasts, but this
is not really the case. That is because these differences
are found in rather obscure organs and external features,
such as amphids and phasmids, that are often difficult for
the non-specialist to observe or appreciate. Nonetheless,
this approach tends to minimize the number of new species
described. It may also severely undercount species if those
species are separated by different criteria.
On the other hand, perhaps the most discerning of nematologists,
the worms themselves, are not bound by such obvious cues and
may discern much more subtle differences. If this is true,
there may be dozens of species within what appears as a single
"morphological" species. This is a situation that
occurs in many phyla of marine animals, and it should be expected
in nematodes. Knowledge of various species' biological interactions
could help to distinguish such cryptic species. Unfortunately,
this knowledge is lacking.
Nematodes occur everywhere and are among the most ecologically
important animals in EVERY ecosystem - including ourselves
and our captive reef ecosystems. Some are free-living, but
as with the flatworms, the vast majority of species are parasitic.
The free-living forms are generally pretty small, ranging
in size from 1 mm to about 20 mm (0.04 to 0.8 inches); however,
they are often exceptionally abundant. In many terrestrial
areas they range in numbers upwards of several billion animals
per acre. Although many parasites are small, most are between
0.5 mm and 60 cm (0.2 to 24 inches) in length, but the largest
nematode is in the species Placentonema, which is parasitic
on the placentas of sperm whales, and reaches lengths of about
9 m (30 feet). Interestingly, and probably because it is so
large (and difficult to study; feature, if you will, the problems
of finding an animal that lives in the placentas of sperm
whales. Step number 1, find a pregnant sperm whale. Step number
2…), this species has become the substance of fable;
it is hard to find any consistent information about it. For
example, its diameter is variously listed in different references
as being anywhere from 0.03 to 2.5 cm (from 1/8th to about
1 inch). I suspect the former value is more likely than the
latter. Females are also reputed to have 32 ovaries.
Every living thing on the planet, with the possible exception
of bacteria, other nematodes and some protists such as amoebas,
is parasitized by nematodes. For example, some nematodes are:
-
totally free-living,
-
free-living as juveniles and parasitic as adults,
-
parasitic as juveniles and free-living as adults,
-
totally parasitic, but with each life stage in a different
host or,
-
totally parasitic with each life stage in the same host.
Then, add to this the fact that some are parasitic in
plants at one stage, and in animals during another.
Thus, if an estimated 10,000,000 animal species are on the
planet (exclusive of nematodes) and if each has at least one
unique nematode parasite (a conservative number, humans have
many...), then there are at LEAST as many nematode species
as all other animal species combined. And this doesn't include
those that are parasitic in more than one host, or in plants,
or the free-living forms.
I should point out as well that you, as you read this, have
nematodes living in your tissues and probably in your gut.
Such knowledge gives a whole new meaning to the quote from
Queen Victoria, who upon hearing of some foible of a cabinet
minister, replied, "We are not amused." While I
presume she could speak for herself, I rather think her worms
could form their own opinions.
Nematodes On and In Reefs
Free-living nematodes occupy just
about every ecological niche. Some are predators, some are
herbivores, while others scavenge detritus for nutrients.
All three types are likely to be found commonly in our aquaria.
Judging from the gut contents I have seen in the animals in
my systems, the scavengers predominate, but more observations
need to be made before anything definitive should be concluded.
Predatory nematodes are common in some marine systems where
they eat foraminiferans, clams or other small shelled prey.
Herbivorous nematodes may be found in algal films or around
clumps of filamentous algae. These worms often suck the cellular
contents out of algal cells. In such environments the nematodes
themselves are likely prey for larger organisms such as some
crustaceans. Scavengers are common in live sand beds where
they scrape bacteria off mineral grains or eat pieces of detritus.
In our systems, as in nature, nematodes perform several important
roles, acting as predators, herbivores and scavengers in a
size range that is normally underrepresented in our systems.
They are really larger than most protozoa, but smaller than
most annelid worms. Consequently, they are more important
to our systems than their small size would imply, as they
facilitate the transfer and utilization of nutrients through
a potential bottleneck in the system.
|
|
Figure 6. An unidentified, about 1 cm (0.4 in)
long, nematode collected from one of my aquaria. The
sensory bristles around its mouth are indicated. Nematodes
that are long relative to their girth, such as this
one, tend to coil when removed from their normal habitat
and placed into water. This animal appeared to be eating
microalgae found in my system's deep sand bed.
|
Aquarists can't do much to encourage the growth and well
being of their systems' nematode fauna. Provided with a fine-grained
sand bed, however, the nematodes will normally thrive. Treatment
of the system with some medications, specifically those for
roundworm parasites, should be avoided, as they will seriously
impact the beneficial worms. If you are concerned about a
fish having parasitic nematodes, treat the fish in a quarantine
or hospital tank, not your reef system.
|