|
Deceptive Simplicity
Although a few organisms
in a couple of other small groups are good claimants to the
title, sponges are widely regarded, and justifiably so, as
the simplest animals (Barnes and Harrison, 1991). While it
is tempting to dismiss such simple organisms as some sort
of failure, such a dismissal would be based more on arrogance
than on fact. In one way or another, within their particular
habitat, all organisms have to be able to do the same kinds
of tasks or overcome the same sorts of specific problems.
Learning the different ways that various organisms accomplish
these same tasks is, in a very real sense, the science of
biology. While sponges are very unlike, in just about all
properties, the readers of this column, both the readers and
any sponges in their aquaria must perform the same basic tasks
of life.
-
They must obtain nutrition or food. Without food, life
stops. All other tasks are secondary to this one.
-
They must get rid of wastes. All organisms create poisonous
metabolic byproducts that we term "wastes."
Scientists consider wastes to be specifically the byproducts
of protein metabolism. For some reason, no animal has
been able to extract, or utilize, the energy in the chemical
bonds between nitrogen and hydrogen (N-H bonds). The major
waste product resulting from protein metabolism is ammonia,
NH3, which, in addition to containing
three metabolically useless N-H bonds, is highly reactive
and exceptionally toxic. As a point of order, what comes
out of the anus (or other such structure...) of animals
is undigested food, often partially processed by bacteria.
While not particularly "tasty," this stuff is
generally not particularly toxic, either.
-
They must avoid becoming food for some other organisms.
How organisms avoid predation often is a defining factor
in their natural history.
-
They must move; actually everything, including sessile
animals and plants, must move. If the perfect organism
never moves, eventually something will happen to it because
of its location, and it will die.
-
They must sense and react to their environment. From
its viewpoint, an organism's environment is defined by
its sensory input. Those senses may be VERY different
from our own. It may seem trite, but aquarists - and scientists
- often overlook that what the organism perceives as its
environment may be very different from what we perceive
it to be. It is hard to overstate how important this fact
is to aquarists. When hobbyists acquire a new animal,
they seldom try to take into account what it will be sensing
from its environment and therefore what it will need in
its new home. Humans are animals that are really defined
by our vision. We think of EVERYTHING visually; even blind
folks say, "See you tomorrow." Being the ultimate
in visually "defined" creatures, it is very
hard for any person to perceive and relate to an organism
that senses its environment mostly by the use, for example,
of chemical sensations. Unfortunately, such a failure
on the part of an aquarist often results in problems.
-
Finally, they must reproduce. Reproduction can occur
either sexually or asexually, but in one form or another,
it has to occur.
The ways that these problems are solved varies from organism
to organism, and the sum of such "solutions" is
unique to each organism. Those solutions define and describe
the organisms, but more importantly for hobbyists, they provide
a blueprint for the husbandry of those animals.
The First Filters
By using "molecular
clocks," it has been possible to roughly deduce when
the first organisms that we might call animals appeared in
the evolutionary history of life on Earth. This rather momentous
event probably occurred sometime between 800 and 1000 million
years ago. These were small organisms, and left no fossils,
so any discussion of their morphology is largely speculation.
However, one thing is clear: they were unable to make their
own food by photosynthesis. This lack of photosynthesis is
the primary hallmark of being an animal. To obtain nutrition
the first animals had to eat other organisms, such as bacteria,
or to eat the byproducts of other organisms, such as sugars,
mucus or their corpses.
Other organisms predated animals, of course, and the most
highly developed of these were likely the animal-like protozoans.
Although the name "protozoa" conjures up images
of primitive "animal" life, in fact, these organisms,
such as amoebas,
ciliates
and flagellates,
represent a diverse array of decidedly non-animal life. Presently,
animals are defined as multicellular; that is to say, built
of many smaller component parts called "cells."
Protozoans lack cells. Although such organisms may be thought
of as having but one cell, it probably is better to consider
them as lacking cells altogether. Many protozoans, for example,
have multiple copies of internal cellular structures; they
just seem to lack the internal division into cells. Good aquarium
examples of such intracellular structural multiplicity may
be found in Caulerpa and other similar algae. Each
individual algal "plant" contains several thousand
nuclei,
but these nuclei are not found within individual cells. Consequently,
each individual of Caulerpa may be considered to be
either a single-celled organism with many nuclei, or a multinucleate
organism without cells.
Some protozoans may be "colonial" organisms, having
several almost independent "individuals" attached
to a single stalk or growing from a common base. Colonial
organisms are particularly common within the two groups of
protozoans referred to as the ciliates
and the flagellates.
It is within the flagellates that are found the organisms
that appear to be most similar to what is likely the ancestor
of sponges and, by inference, the ancestor of all animals.
These organisms are called choanoflagellates
(2,
3,
4,
5).
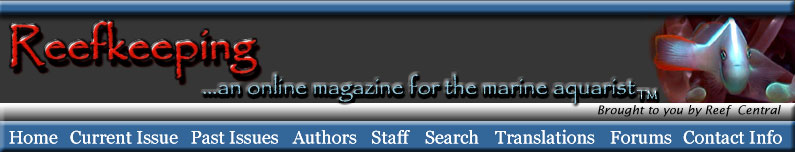
Colonial choanoflagellates are clusters of small, rather spherical
cells, each of which has a structure resembling the top flaring
part of a funnel on its surface. In the center of this funnel
is a single microscopic hair-like flagellum. As the flagellum
beats it creates minute water currents that push water up
its length, out away from the cellular surface. This movement,
in turn, draws water from the sides and directs it in toward
the base of the flagellum. To get there, the water has to
pass through gaps in the wall of the "funnel," which,
in reality, is comprised of tiny finger-like projections referred
to as microvilli. The microvilli trap small particulate materials,
mostly bacteria, and transport them to the cell's surface
where they are ingested. The shape and function of choanoflagellate
cells is virtually identical to that of choanocytes
(2,
3,
4),
the most characteristic type of sponge cells, whose name literally
means "funnel cells." Choanocytes
were first seen in sponges in the early nineteenth century,
and were considered to be unique to, and absolutely characteristic
of, sponges. If you found a choanocyte in some unknown animal,
it had to be a sponge, as no other groups had them. Then,
starting in the late 1970s, cells of similar construction
were found in many other animal groups, including vertebrates.
This presence of choanocyte-like cells throughout much of
the animal kingdom is considered to be one line of evidence
for the common descent of all other animals from an ancestor
containing these cells.
Figure 1. A diagrammatic view of a choanocyte. For
images
of actual choanocytes, follow the links in the text.
The Body of the Beast
Sponges don't have the simplest type of structure found in
animals, but they are close to that limit, and their structure
is neither difficult to discern nor to understand (Harrison
and De Vos, 1991). These are animals that get their nutrition
by filtering water through themselves. If that fact is kept
in mind, their body form becomes quite explicable and reasonable.
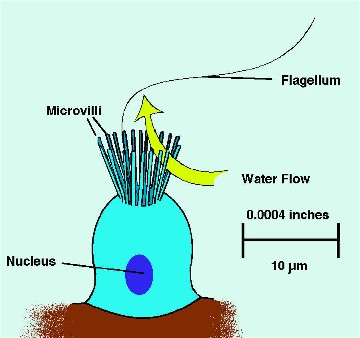
The simplest sponges are tubular animals that pull water through
the sides of their body into the center of the tube and then
blow it out both ends of the tube. Larger sponges have much
more complicated water filtration pathways, but otherwise
the sponges' body structure is relatively simple.
There really is nothing like a sponge throughout the rest
of the animal kingdom. Most sponges, quite literally, suck
water in through tiny holes covering their body's entire surface,
filter it clean of all acceptable foods, and flush it out
through large drainage pipes. Depending on the type of sponge,
the tiny holes that allow water to pass internally may be
so small that they pass through a single cell, or they may
be somewhat larger, but still tiny, openings to small tubes.
These passageways are pipes or conduits constructed of cells.
The water is pulled into the sponge by way of the surface
pores and is moved along inside the water channels by the
beating action of the flagellated choanocytes lining "filtering
chambers." Downstream of the filtering chambers the
water channels get progressively larger until the water leaves
the sponge through a large aperture, called an osculum.
A large sponge may have millions of these small entry pores,
each of which is called an ostium,
and one to several hundred excurrent apertures.
When water is passed through the filtering chambers, it moves
through the fine microvillar comb surrounding the bases of
the choanocyte flagella and particulate materials, bacteria
or phytoplankton get removed from the water and eaten by the
choanocytes. When they have eaten enough food, the choanocytes
are able to perform one of the better metamorphic feats in
the animal kingdom. They may "reel in" their flagellum
and turn into an amoeboid cell, called an archeocyte. These
cells can wander all over and through the sponge. Archeocytes
are totipotent
and may perform any task within the sponge with the
exception of becoming a gamete. Many different functional
cell types have been described within sponges; these have
been called choanocytes (filtering cells), myocytes (contractile
cells), porocytes (cells having a pore in them) and pinacocytes
(surface lining cell), to name but a few. Such names are illusory,
however; the cells are named by their function at the moment
of observation. In reality, all of them can revert to being
an archeocyte and wander away to become something else.
Figure 2. A diagrammatic representation of a simple
sponge showing the basic
features of sponge anatomy.
Given the mobility of the cells that constitute the living
"stuff" of sponges, it is not surprising that sponges
lack tissues. And, lacking tissues, they have neither organs
nor organ systems. Most sponges can be thought of as a group
of cells rather loosely working together. Unlike all other
animals, in the sponges, the constituent cells are only casually
connected to each other; in fact, with vigorous action some
sponges can literally be shaken apart resulting in a slushy
mixture of living cells and mesogleal components. "Sponge
smoothie, anyone?" This can occur because sponges lack
what biologists call "tight junctions" between the
cells. Tight junctions are really the "glue" of
animal life; they are fine molecular threads connecting the
cell membranes of adjacent cells. In all animals except sponges,
the cells ALWAYS are securely glued together by tight junctions.
No other type of animal can simply be shaken apart; its cells
would tear and be destroyed before they would separate.
Sponge cells are typically arranged in outer and inner layers
living over, and lining, a middle skeletal layer called the
mesoglea. The term mesoglea literally means "middle glue"
and that name for the middle region of sponge's body wall
approximates its function. The mesoglea is not sticky like
any adhesive, but rather consists of the sponge's skeletal
material, mostly spicules
or protein, and is largely formed by the secretions
of the cells surrounding it as well as of some cells that
wander around within it. While not without living cells, the
mesoglea doesn't have many cells in it relative to the number
of cells in the sponge's inner and outer surface layers. The
reproductive cells also generally reside within the mesoglea;
presumably they get more protection in that location than
they would on the body's surface.
The Ultimate in Fragging
One of the classical demonstrations of this "fragility"
of sponge structure can be done by aquarists. You will need:
-
A clean glass container, something never washed by soap;
soap and soap films are lethal to most marine life.
-
Some good clean seawater. Artificial seawater may work,
but real sea water filtered through a one micrometer filter
is better.
-
A blender with clean blades and container, see the comments
about soap above.
-
A coarse (around ¼ mm) mesh screen or cheesecloth.
-
Two different colors of sponges from your aquarium.
Fill the blender with about one cup (250 ml) of seawater.
Add two small (1 cm x 1 cm x 1 cm or smaller) pieces of the
two sponges. Turn the blender on and let it rip until you
have a "sponge shake." Filter the mess through the
coarse screen or cheesecloth to remove all visible chunks.
Pour the remaining "juice and goodies" into your
clean glass container. Cover the container loosely, then put
it on a shelf and don't disturb it. Mark the water level with
a pen and make sure that evaporation is replaced with fresh
distilled or RO/DI water daily. Maintain it at tank temperature,
if possible; if not, any temperature in the middle to upper
70°F range will likely work. After a few days small globs
the same color as the original sponges may become visible.
Some of these will move, amoeba-fashion, to find and fuse
with other globs of their own color. These small masses are
tiny sponges reconstituting themselves from the surviving
cells of the blenderized sponges. Interestingly enough, though,
cells from one sponge will be able to "recognize"
other cells from their same sponge and will not fuse with
cells from the other sponge.
Reproduction
Given the ease with which most sponges can recover from injury,
it is not surprising that these animals are masters of asexual
reproduction. Demosponges, particularly, grow well from fragments
and in some areas clonal sponge populations contain many separate
individuals resulting from widespread asexual reproduction
due to fragmentation (Hartman and Reiswig, 1973; Reiswig,
1983). Additionally, a few marine sponges and many freshwater
sponges produce asexual "resting bodies." These
structures contain numerous archeocytes in a state of dormancy,
and often they are surrounded by a resistant outer shell or
coating. When the conditions change or improve, the coat ruptures
and the cells within can differentiate into a small sponge.
Sponges also reproduce sexually. In general, both eggs and
spermatozoa are produced in the mesoglea. During spawning,
the sperm generally are released as a dense cloud of "milky"
water blown out of the osculum. Ova generally are retained
within the mesoglea. In many cases, fertilization is rather
complicated. The sperm is caught by a choanocyte of a female
sponge and ingested. The sperm's nucleus, containing its genetic
material, is encased in a membrane, and the choanocyte changes
into a "carrier cell" which then takes the form
of an amoeba and moves though the mesoglea to find an egg
and deliver the sperm's nucleus to it. Embryonic development
occurs within the mesoglea until a relatively large flagellated
larva is formed. Given the diversity of form within the various
sponge groups, it is not surprising that several different
types of sponge larvae have been described. The larva escapes
or is released from the parent and swims away. Sponge larvae
typically do not feed, but swim around for a while until they
choose a site upon which to settle, fasten to the bottom and
metamorphose.
Except for slight contractions of tubular osculae, the movement
that occurs as a larva is the sum total of movement that most
sponges are capable of, particularly sponges that reach relatively
large sizes. Some smaller sponges, however, including several
species of Tethya that are commonly found in aquaria,
are quite mobile and capable of both changing their shape
and moving at rates of several centimeters per day across
acceptable substrates. (See movies
of sponge movement.) The mechanism by which such movement
occurs is not yet completely understood, but it appears to
be the result of a sort of amoeboid movement by the sponge's
basal cells in contact with the substrate.
Spongy Thoughts
One of the more interesting things that
has happened over the last ten years or so has been a change
in the "appreciation" of sponges. We used to think
of all possible spongy animals as being both similar in structure
and closely-related; such a view is reflected in their treatment
in older invertebrate zoology textbooks, such as those by
Kozloff (1990) and Ruppert and Barnes (1994). Genetic investigation
has changed that viewpoint rather significantly, and has discovered
that modern sponges are probably three distinct groups arising
at different times and relatively distantly related to one
another. In other words, the concept of the phylum "Porifera"
as a taxonomic unit containing all the sponges, and descended
from a common ancestor, is no longer supported. It now appears
that the three groups previously considered the major taxonomic
subdivisions or classes of the phylum Porifera should each
be considered to be distinct, and a group unto itself (Halanych,
2004). As the groups are each distinct and are each descended
from a common ancestor, there should be no problem assigning
each to its own phylum. Presently the discipline of animal
taxonomy, however, is undergoing considerable flux, and the
whole concept of a phylum, or something like it, is changing.
For the moment, it seems prudent to say that three distinct
living types of animals may be called sponges and that at
a basic level they may be easily distinguished by the type
of skeleton they possess (Ruppert et al, 2003). In
addition to variations in skeletal composition, numerous other
characteristics also separate these groups. Those characteristics'
differences often are obscure and require microscopic examination,
so it seems easiest for this discussion to define the types
of sponges by their skeletons. These groups are:
-
The so-called "Glass Sponges," in the group
Hexactinellida, characterized by a skeleton comprised
largely of fused silica;
-
The "Regular Sponges," in the group Demospongiae,
characterized by a skeleton comprised of some combination
of silica spicules, protein fibers and, in some forms,
calcareous masses, and
-
The "Calcareous Sponges," in the group Calcarea,
characterized by a skeleton comprised of calcareous spicules.
Hexactinellida
The hexactinellids are the most
unusual sponges in a lot of ways. Except for their gametes,
they lack cells. Instead of cells, their skeleton of fused
spicules is covered by a thin protoplasmic mass that is elaborated
into cellular-like structures, but few, if any, cell membranes
separate or delineate these structures. Such an acellular
multinucleate mass is termed a "syncytium."
Syncytia are common among smaller invertebrates; for example,
most tissues or organs in rotifers or roundworms don't have
cells, either. However, in these groups, these syncytia often
are considered to be adaptations for their small size, the
idea being that in an animal the size of a rotifer, cell membranes
literally take up too much space. Hexactinellid sponges, however,
are often large; presumably, they have developed syncytia
for other reasons. The mesogleal layer is reduced to a skeleton
of fused spicules and free spicules imbedded in the syncytial
mass. The skeleton may be composed of spicules of various
shapes, but spicules with six rays (called hexacts) predominate.
Typically, many such spicules are fused together to form a
brittle and inflexible skeleton. Relatively few animals have
the capability to metabolize and secrete silicon dioxide,
but sponges do and the hexactinellids are masters of this
art. The fusion of different spicules, which is done inside
the syncytial mass, results in a relatively strong, solid
structural mass. Many of the hexactinellid sponges are asymmetrical,
but others may be cylindrical with the spicules arranged in
beautiful geometric patterns.
|
Figure 3. The "cloud sponge" from the
Northeastern Pacific, Aphrocallistes vastus.
This is one of the few hexactinellids found in shallow
water. The animal pictured here was about 1 m (3.3 ft)
high and 2 m (6.6 feet) across. These animals are common
from the Gulf of Alaska south through British Columbia
at depths of about 30 m (100 ft) or more.
|
Glass sponges are typically found in deep waters and are,
in some instances, characteristic of the deep seas. Many of
them tend to be relatively large animals; individual sponges
are commonly more than a meter (3.3 ft) in height or diameter.
They are immobile as adults; their only mobile form is their
larval form. They have no sensory organs or structures; however,
their entire syncytial surface probably is sensitive to various
chemical and tactile stimuli.
Little is known about the natural history and ecological
relationships of most hexactinellids, but some work has recently
been done, mostly on temperate forms (Leys and Lauzon, 1998).
They tend to be long-lived, slow growing animals, and the
ages of some have been determined to be in excess of 200 years.
They are relatively uncommon on coral reefs, and it is unlikely
that any reef aquarist would encounter one. Many of the basic
questions about them remain to be answered; for example, in
many cases we do not know what their predators are, or how
they are protected from predation, although the fact that
their body contains myriads of spicules that are really nothing
more than shards of sharp glass is presumed to have something
to do with the fact that most predators seem to avoid them.
It is likely that they are chemically protected against predation
as well.
Demospongiae
The majority of sponges are demosponges. As do the hexactinellids,
they typically have spicules made of silica, but the spicules
are never fused to form a solid lattice. Nevertheless, the
spicules are often cemented together with proteins into a
network that may be as complex as that found in glass sponges,
if not as permanent. As befits a group containing thousands
of species, there is a lot of diversity of skeletal structure.
A few demosponges lack spicules altogether and have only a
proteinaceous skeleton; these are the classic "bath sponges."
Some others, called "sclerosponges," secrete a massive
calcareous skeleton, in which the silicate spicules are imbedded.
A thin tissue layer overlies this massive skeleton. Fossils
that appear to be very similar to such sponges are fairly
common in the fossil record from the mid-Paleozoic, when they
were reef-forming animals. While predominantly a marine group,
the Demonspongiae also contains the only sponges found in
freshwater. Freshwater sponges are not very diverse, but they
are very common, being found in most non-polluted freshwater
ecosystems.
Figure 4. Large demosponges, such as this Niphates
digitalis, are
common in Caribbean coral reefs.
Demosponges tend to be moderately-sized animals, but some
are found growing as only thin layers over rocks. In contrast,
others may exceed a meter in height. In many temperate nutrient-rich
areas and deep-water coral reef areas (Suchanek, et al,
1983), demosponges are the dominant benthic animals. These
sponges tend to grow more rapidly than hexactinellids, and
it is not surprising that they are commonly found in virtually
all coral reef habitats, and hitchhike into reef tanks in
or on live rock. In general, those sponges found living in
crevices, within rocks or in lagoons tend to do best in reef
aquaria. Those demoponges that require a lot of currents,
such as the brightly colored so-called "tree sponges"
and "ball sponges," generally perish after a short
period in reef tanks.
Many animals eat demosponges, but on coral
reefs their primary predators are fishes, various snails,
such as nudibranchs, and sea stars. Having so many predators,
natural selection has casued the evolution of the sponges
either to hide or to be very toxic. Generally, the large,
evident sponges on coral reefs appear to contain toxic chemicals.
These sponges are often long-lived and many of them are homes
to other animals that live on or in them in various symbiotic
relationships. Many of these animals are commensals, which
benefit from living on the sponge, but whose presence doesn't
benefit the sponge. Others appear to be ectoparasites, intercepting
and eating foods brought to the sponge by its filtering currents.
Still others may benefit the sponge in some manner.
Lacking structures for attacking other animals, sponges might
seem to be at a disadvantage in the rough and tumble competitive
world that constitutes a coral reef's benthic environment.
This is decidedly not the case, however. Sponges are powerhouses
of chemical synthesis, and many produce highly toxic chemicals.
These may serve to make them unpalatable to predators, but
similar (or the same) chemicals also may be liberated from
the sponge to kill any nearby animals. On a natural reef,
where water exchanges are continuous, these chemicals generally
act over only very short distances, from a few millimeters
to a centimeter or two. In an aquarium, the water movement
is not sufficient to flush the chemicals from the system.
In such enclosed environments, highly toxic sponges may well
have the potential either to kill potential competitors, such
as corals, or to stress them so severely that they become
diseased and die. Unfortunately, while it is clear that many
sponges may be able to do this, there is no practical way
to determine if such chemical releases are occurring and what
their affect is; however, it is beyond neither belief nor
reality that sponges are responsible for many of the mysterious
deaths that plague aquarists.
Calcarea
Sponges with skeletal spicules made of calcium carbonate
form the third group of sponges, which contains a few hundred
species. Some calcareous sponges are commonly found in many
reef aquaria. These sponges tend to be small animals, seldom
reaching 15 cm, about 6 inches in any dimension. They also
tend to be simpler in structure than the demosponges and hexactinellids.
Figure 5. Calcareous sponges, such as these 2 cm (0.8
inch)
high individuals of a species of Leucandra, tend to
be
small and drably colored.
Reef Aquarium Sponges
Sponges are normal and common components of coral reef ecosystems,
but may or may not be good things to have in reef aquaria.
Many sponges are good competitors and can crowd out more "desirable"
livestock; additionally, many of them are highly toxic. These
properties, coupled with the difficulty in identification,
make sponge husbandry a most interesting topic, and one I
will discuss in detail in next month's column.
Figure 6. Two coral reef sponges spawning. These are
males releasing a sperm suspension in the water currents leaving
the sponge through the osculum. The females retain the eggs
and fertilization will occur internally. Images courtesy of,
and with thanks to, Eric Borneman.
|

)


)
)
