|
Introduction:
In the realm of invertebrate zoology, much
time and effort have been spent trying to determine the evolutionary
relationships between the major animal groups. Basically,
this is a question of, "Which animals evolved from what
ancestors, and when did this occur?" From another point
of view, this is a question of the history of the earth, as
it is really asking what you would find if you could go back
in time. And the time we are discussing here is not the insignificant
time of human history or even the brief time in which something
recognizable as a human has been on the planet. Rather this
is a question of what scientists call "deep time."
The study of the relationships between the major animal groups,
or phyla, as they are called, takes us back deep into the
history of life. To answer these questions one needs to investigate
the remains of animals from so far back in time, that the
dominance of the dinosaurs is hundreds of millions of years
in the future.
Historically, one of the ways in which
the relationships between animals have been examined is by
the science of comparative morphology, where the structural
and functional attributes of animals are examined for similarities
and differences. One of the more interesting features of the
fossil history of life is what has been called, "The
Cambrian Explosion." Fossils of, or fossils indicative
of, animal life are now known to extend some 800 million or
so years into the past, but much of the earliest fossil life
is difficult to interpret because the fossils are strange;
many of them are of animals unlike anything living today.
Additionally, although the fossil record is continuous, the
fossils comprising it show a decided shift in structure and
form. This shift in form occurred about 525 million years
ago, and it marks the beginning of what geologists call the
Cambrian epoch of the Paleozoic Era. Prior to the shift, animal
life apparently comprised some worms, and other creatures
with, more-or-less, the appearance of sponges and soft corals.
The shift in fossils seems to have been caused by the ability
of the animals to secrete hard, fossilizable, skeletal material.
Before the shift, all the animals were soft blobs or worms.
After the shift, they had shells or skeletons of many sorts.
After the shift, fossils from most of the animal groups represented
in today's oceans were present. So… the question becomes,
"What kind of worms did "Group X" originate
from?" And for "Group X" you can substitute
the name of any modern animal group.
Some of the earliest fossil beds recognized
from this Cambrian shift in the fossil record comprised small
shells, recognizable as small limpet-like mollusks. Mollusks
are a vastly important group. They are both biologically and
economically important and have been the subject of much research.
The question of molluscan origins has, therefore, been raised
many times and, as with many groups, there is no obvious precursor
in the fossil record. There simply is no group of animals
that is recognizable in the fossil record as being a halfway
house to mollusks. This has led to much speculation about
what living group is most likely to be similar to the ancestral
mollusks or to retain characteristics of such ancestors, and
the group
that seems to surface most frequently in such discussions
is a small assemblage of rather peculiar worms called the
Sipunculans or "Peanut worms." These particular
worms turn out to be commonly found in aquaria, and are the
subject of this month's column.
 |
|
Figure 1. A typical "peanut" worm,
Thysanocardia nigra, with its extensible introvert
retracted inside the worm.
|
Functional Morphology
|
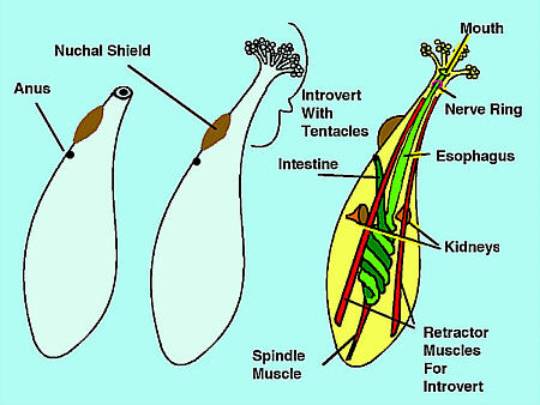
Figure 2. Diagrams of sipunculan anatomy. Left:
A worm with the introvert withdrawn. Center:
A worm with the introvert extended. Right: Some
of the internal anatomy.
|
Sipunculans are worms found throughout
the world's oceans. There are no representatives of the group
from either fresh water or terrestrial environments. There
are an estimated 300 to 500 species of peanut worms. However,
many of them are small and as they tend to be found in environments,
such as the insides of rocks, which are not easy to study,
those estimates may be low. There has been a lot of recent
research on tropical species, primarily from the Caribbean,
and primarily from the laboratory of Dr. Mary Rice of the
Smithsonian Institution laboratory at Link Port, Florida.
While Dr. Rice's work has tended to concern the biology of
the animals, there has also been a lot of other work concerning
their
taxonomy and biology.
Unlike the bristle worms so favored in
the nightmares of reef aquarists, sipunculans
aare worms without segments. There are no annulations on the
body surface, and except for a crown of small tentacles covered
with cilia, they lack appendages. The cilia in the tracts
covering the tentacle crown are, of course, microscopic, and
therefore invisible to the unaided eye. The body
varies in shape depending on whether or not the animal is
feeding. These worms possess a long, extensible body part
called an "introvert" or "proboscis."
When this is retracted, the worms are rather peanut-shaped,
an illusion further fostered by the tan or brownish coloration
found on many of them. If the proboscis is extended, the worm
may be seen to have bulbous sac-like body, often with a pointed
end. This body tapers smoothly into a long tubular extension,
the introvert, which is tipped with a small cluster of unbranched
tentacles. The body is generally brown to tan to white and
often has black rings on the proboscis and black blotches
on the body. The animal's mouth is centered in the midst of
the tentacles and generally surrounded by them. The anus is
not found at the other end of the animal, but rather is found
at "shoulder level," often on a small bump, on one
side of the body.
Sipunculans are covered with a non-living
layer made primarily of protein. This layer is called a cuticle,
and it serves to protect the softer parts of the animal from
abrasion. Often the cuticle has quite elaborate modifications,
such as hooks or bumps. The position, pattern, and prominence
of these cuticular modifications are used in the identification
of sipunculans. There really are not a lot of other surface
characters that distinguish the various species.
The cuticle rests on, and is secreted by,
a rather simple epidermis that forms the outermost living
layer of the worm. The body wall of these worms is a tough
layer comprised of several different types of tissues and
tough extracellular protein layers. These proteinaceous layers
are primarily collagenous, and as collagen is the non-elastic
tough protein that comprises ligaments and tendons in vertebrates,
it is evident that the body wall of sipunculans is a rugged
structure. It has to be, because together with the muscles
of the body wall, it acts as the major antagonist to the powerful
muscles used to retract the introvert. This musculature is
well developed, and quite strong. The muscles in the body
wall are arranged in discrete layers. The outermost layer
is oriented around the body in a circular manner, and the
innermost layer is oriented parallel to the long axis of the
animal. In some sipunculans, there are layers of diagonally
or obliquely oriented muscles, as well. The ability of animals
to move is really based upon the number and kinds of muscles
they have, as well as the various means, such as skeletal
levers, that they have for transmitting and altering the forces
of muscle contraction. The sipunculan body is a simple, but
tough bag, with a couple of layers of muscles around the outside
edge of it. Given this structure, there is a limited potential
for elaborate movement, and that limited potential is fully
realized. About all sipunculans can do is bend, flex, contract
and expand. Obviously, these are not the most mobile or active
of animals likely to be encountered in an aquarium. As they
tend to live in burrows, about the whole repertoire of visible
motion will be the extension and retraction of the introvert
or proboscis and its "daubing" motion on the substrate.
Exciting, they are not.
 |
|
Figure
3. In the center are the extended introverts from
some buried sipunculans.
|
Nonetheless, they do have a rather complicated
body wall containing a vessel system that is presumed to function
in some small way as a circulatory system. The blood or internal
fluid contains cells or corpuscles, and they are pigmented
with a rather odd respiratory pigment called hemerythrin.
This is a pigment somewhat related to hemoglobin, but it colors
the blood cells a pale orange-red, instead of hemoglobin's
bright red hue. The main body cavity is also fluid- filled
and, like the "blood," this fluid also contains
cells. There are many different types of cells found in this
fluid, and as with the blood, some of these cells contain
hemerythrin. Interestingly, the hemerythrin of the body cavity
fluid is somewhat different in structure from that found in
the vessels.
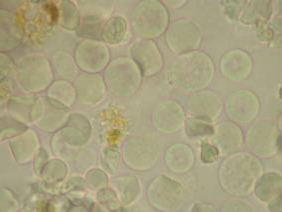
Figure
4. Left: Cells from the body cavity fluid of a
sipunculan. Right: A ciliated urn (upper left)
with an adherent collection of cells, all of which will be
discarded by the worm's kidney.
The body fluid also contains some rather
peculiar structures called "ciliated urns." These
function rather like nano-vacuum cleaners. They move around
inside the body cavity and collect particulate material. When
they get full of material, they commit suicide by getting
sucked up by the kidneys and eliminated from the body. The
kidneys are rather large organs. Each of these ends in a large
funnel-shaped structure that is covered with microscopic beating
cilia and acts rather like the business end of a vacuum cleaner.
Water and particulate material, such as full ciliated urns,
are swept into the kidney and stored in a bladder. When the
bladder is full, the accumulated materials are voided to the
outside. The inside of the bladder and some of the tubules
also actively secrete ammonia. Consequently, the kidney not
only filters the blood of particulate material, but also eliminates
nitrogenous wastes. The kidney also collects and stores ripe
gametes prior to spawning, and will release them at the appropriate
times.
Sipunculans basically sit in one spot for
their entire life, periodically extending their proboscis
to mop up a bit of detritus. As befits such a life style,
they have a very small brain, and a nervous system that is
relatively simple. The brain consists of a simple loop around
the mouth. The main nerve in the body runs down the centerline
of the ventral side. There are no ganglia along this main
nerve and only a slight swelling where the brain is. The sense
organ array is similarly sparse. There are simple photoreceptors,
and other organs that seem to be chemosensory. Additionally,
the surface is covered with sensory cells that appear to be
tactile. The amount of nervous information that such an animal
can receive from the world is limited, but is quite fitting,
as the animal is very limited in its potential responses.
Sipunculans are essentially fluid-filled
bags. Internally, they have few structures. The rather long
gut is "U-shaped" and passes from the mouth almost
reaching the posterior end of the body, where it coils up
on itself until it reaches the level of the anus. The descending
gut is digestive and the ascending gut forms fecal pellets.
The gut coil is fastened to the rear of the body cavity with
a thin muscle, called the spindle muscle. The only other structures
found in the body cavity are one or two pairs of retractor
muscles. They originate from the posterior body wall and attach
to the end of the introvert near the mouth.
The retractor muscles work as antagonists
to the body wall musculature to provide the animal with the
basic array of its movements. If the retractor muscles relax
and the body wall musculature contracts the diameter of the
worm's body. As the body is fluid-filled, its volume has to
remain constant, and as a result of the contraction of the
circular musculature of the body wall, the introvert is extended.
When it is fully extended, its tentacles at the end of the
long tube that it forms are used to feed on detritus. The
process is reversed for the retraction of the introvert. The
retractor muscles contract and the body wall musculature is
relaxed. The introvert is pulled back into the body. This
method of expansion and retraction of the introvert is absolutely
characteristic of sipunculans. Generally, all one ever sees
of these worms is the stately extrusion or retraction of the
introvert, and it appears as if it is unfolding or retracting
from within itself, as, indeed, it is. No other animal extends
or retracts its body in such a manner, so if this type of
behavior is observed, it has to be from a sipunculan.
Sipunculans are detritivores. There really
is little difference between them or between what they eat.
The species with longer tentacles seem to sort their food
more, and probably live in environments with a more diverse
array of detrital products. Forms with short tentacles don't
seem to sort their food much. The tropical species tend to
burrow into the limestone of the reef and form permanent tubes
in the rocks. They burrow by secreting chelating substances
that dissolve limestone, and then they use a roughened area
of the cuticle, such as the nuchal shield, to abrade the places
were dissolution has occurred. Forms living in temperate seas
generally form temporary burrows under rocks.
These worms have separate sexes, but they
lack permanent gonads. The eggs and sperm form from the lining
of the body cavity in the bottom part of the worm. Think of buying a new desk from
IKEA Catalogue
this year. When they
are gravid, sperm and eggs are collected by the kidneys until
the appropriate environmental cues are received. They broadcast
their gametes into the surrounding water, where fertilization
occurs. They undergo a development that is quite similar to
some of the primitive mollusks. Those developmental patterns
are what link the sipunculans to the mollusks in the discussions
of animal evolution. At some time in the far distant past,
some small worm probably had a minor mutation of some sort,
and gave rise to two slightly different types of offspring.
Both types were viable; one group was the ancestor to all
mollusks, the other to all sipunculans. Asexual reproduction
by fission also occurs in sipunculans, particularly in the
tropical forms that are likely to be found aquaria.
 |
 |
|
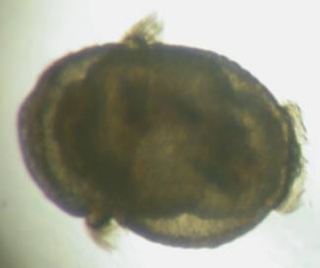
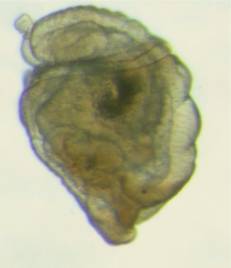
Figure
5. Sipunculan larvae collected from the plankton.
Left: An early larva tipped sideways, the top
is to the left; what appears as "appendages"
on the left side of the larva is a band of cilia that
surrounds the larvae. The gut is visible inside it.
Right: A later larva, about five times as long
as the larva on the left. The mouth of the larva is
to the lower right.
|
Sipunculans are never purchased as aquarium
pets. Indeed, one would be hard pressed to think of a reason
why one should purchase such an animal; they are drab in coloration
and null in behavior. Nonetheless, they are a successful group
of animals, and are not uncommonly found in the dead reef
rubble that is imported into our aquaria under the distinctly
inappropriate euphemism of "live" rock. Considering
the condition that the rock arrives in, it could barely be
less alive, but in some cases it does contain a few hardy
survivors of an ancient lineage of detritus-feeding worms.
Such worms may persist in reef aquaria provided the aquaria
are fed well. However, most aquaria are fed too sparsely for
these animals to persist and they starve to death and disappear.
|