|
Introduzione
Questo mese avevo intenzione di scrivere
la seconda parte dell'articolo sull'allevamento dei coralli
Domenicano, ma non èro di stesso umore per continuare
il tono "divertente" della prima parte di quell'articolo.
Quindi, piuttosto di sforzarmi, ho deciso di cominciare un
articolo in due parti che sto progettando gia da alcuni anni.
Da anni, accumulo rapporti del rilascio delle uova dei coralli
in acquario. Questo per me e una delle ultime mete in allevamento
dei coralli in cattività. Ho gia scritto tante volte
che il nostro successo in allevamento di coralli sarà
misurato, in gran parte, dal numero di prevedibili avvenimenti
di riproduzione sessuale. Fino ad un tempo fa, le uova coralline
in acquario erano eventi sporadici e senza alcuna regolarità
o periodicità. Quest'ancora è in gran parte
vero, ma i casi stanno diventando più frequenti, specialmente
nelle vasche relativamente nuove, curate dalle persone senza
l'esperienza negli acquari di barriera. A me, questo è
importante.
Finalmente, e forse più importante,
per almeno alcuni degli eventi di rilascio delle uova, che
sono stati osservati e fotografati in acquari, e che sono
una testimonianza d'eventi veramente originali.Come primo
sì e vista la riproduzione di una specie, quale non
è stata mai testimoniata o documentata neanche in natura.Questo
serve come un'importante osservazione per la comprensione
generale della natura di riproduzione sessuale di quella specie.
Secondo, per almeno alcuna specie che ha deposto le uova in
acquari, il metodo di riproduzione sembra essere completamente
diverso di quello che è stato osservato o è
stato documentato in natura. Questo è, a dire il minimo,
molto interessante.
Comincerò questo mese con una spiegazione
generale della riproduzione sessuale di coralli, e continuerò
il mese prossimo con un documento molto lungo pieno di fotografie
d'eventi del rilascio delle uova di coralli in diversi acquari.
Mostrerò anche come queste uova sono simili o diverse
di quelle conosciute in natura.
Riproduzione sessuale sulle barriere corallines
Riproduzione e nuove crescite dei coralli
sulle barriere coralline, sono dei processi più critici
che permettono a loro di rimanere presenti in tutti i periodi
geologici. Coralli si riproducono sessualmente ed asessualmente.
Molti coralli sono capaci di spargersi sulla barriera asessualmente,
per esempio con frammentazione, ma questo dà luogo
a cloni della stessa colonia e non aumenta la diversità
o risorse genetiche della barriera. Se tutti i coralli si
spargessero con frammentazione o altro modo asessuato, le
barriere hanno potuto cessare di esistere gia da qualche tempo,
o almeno èssere molto diverse, forse vittime di una
crisi d'inincroci (accoppiamento tra soggetti consanguinei),
con alcune specie monospecifiche, che competendo l'una con
l'altra, eventualmente realizzarebero il dominio sulle barriere.
Riproduzione sessuale richiede la fusione
di gameti maschili e femminili (sperma ed uovo). Questa fusione
forma uno zigote che cresce in un embrione, che alla fine
si sviluppa in una larva che nuota libera ed e chiamata planula.
Uova e sperma sono prodotti nelle accumulazioni di tessuto
gastrodermale (il più interno dei due strati di tessuto
corallino) sul mesentere, rispettivamente chiamate ovaie e
gonadi. Planule sono estremamente diverse tra le specie,
ed alcune hanno la capacità di esistere nel plancton
per periodi lunghi (più di 100 giorni in alcune specie).
Questo lascia la possibilità di dispersioni molto lontane
e l'abilità potenziale di coralli di seminare lontano
dalla colonia d'origine. Riproducendosi sessualmente, si acquisisce
la diversità da: 1) i contributi delle celle sessuali
maschili e femminili delle colonie; 2) l'eventi di incroci
meiotici durante la formazione di celle sessuali; e 3)l'aumento
delle risorsa genetica avendo colonie nuove arrivate dal di
fuori della comunità locale attraverso la dispersione
di planule. Questo ha implicazioni pratiche sulle barriere
che sarebbero perse dalle catastrofi naturali o artificiali.Loro
sopravivrebbero, in che la stessa risorsa genetica probabilmente
esisterebbe in un luogo lontano, e che con tempo, potrebbe
riseminare la barriera che è stata distrutta.
 |
Il "Orientamento Sessuale" di Coralli
No, questo non è uno scherzo che
esige correttezza politica. Coralli possono essere divisi
in due gruppi maggiori in riguardo alla loro natura sessuale.
Il primo gruppo è diviso in due
sottogruppi che si distinguono se o no una colonia produce
sperma ed uova separate. La specie "Gonochorica"
(anche nota come "dioecious"), o quegli in che colonie
sono maschi (produttori di sperma) o femmina (produttrici
delle uova), comprenda approssimativamente 25 percento di
coralli studiati. I rimanere 75 percento sono considerati
ermafroditi, dove una sola colonia produce sperma ed uova.
Il termine ermafrodito è interessante; viene dalla
mitologia greca. Hermes era il messaggero degli dei, ed Aphrodite
era la dea della bellezza. Quindi, il "messaggero"
di sperma maschio consegna il suo pacco all'uovo femmina "dea
di bellezza". Sei specie, Agarcia agaraciles,
A. humilis, Galaxea fascicularis, G. astreata,
Caryophyllia ambrosia, e Porites astreoides
sono state riportate come specie con il potenziale per essere
gonochoriche o ermafrodite.
Ermafroditismo può essere suddiviso
ulteriormente in tre altri gruppi. Nel primo, "ermafroditi
simultanei", sperma ed uova sono prodotti contemporaneamente.
L'altra specie è, colonia maschile funzionale prima,
e poi si sviluppa in colonia femminile funzionale. Questo
e stato chiamato "protandry". D'altra parte esistono
le colonie femminili funzionali prima, e poi si sviluppano
in colonie maschili funzionali. Questo e stato chiamato "protogyny".
Specie che è o protandrous o protogynous, e stata chiamata
"ermafroditi sequenziali". La maggioranza di coralli
ermafroditi ed ermafrodita simultaneo.
Il secondo gruppo è diviso in due
sottogruppi che dipendono come i gameti sono entrati uno con
l'altro in contatto. "Broadcast spawners " rilasciano
contemporaneamente uova e sperma nella colonna d'acqua per
la fertilizzazione esterna e sviluppo successivo ."Brooders"
hanno uova fertilizzate internamente con sviluppo del planule
all'interno dei polipi. La maggioranza di coralli studiata
è spawners (approssimativamente 85 percento), col rimanere
15 percento brooders. Fino un tempo fa si credeva, che la
maggior parte di coralli era "viviparo", o brooders.
Notevolmente contrari agli altri animali, alcune specie coralline,
come per esempio Pocillopora damicornis, esibiscono
entrambi metodi riproduttivi in ubicazioni diverse. Si è
proposto anche che questo può essere una ragione valida
di esaminare tali popolazioni per vedere se loro sono specie
veramente separata o una sottospecie.
L'Importanza d'Essere Diverso
Le variazioni in riproduzione sessuale
in coralli sono teoreticamente importanti in diversi modi.
Per capire meglio l'importanza di questi aspetti, si può
affermare che ogni metodo ha le varie caratteristiche che
possono essere applicate alla specie generalmente, o comunità
che li utilizzano. In termini di una specie ermafrodita o
gonochorica, ermafroditismo è favorevole nelle piccole
popolazioni perché così là probabilità
di avere la fertilizzazione riuscita aumenta. Anche un ermafrodito
simultaneo può produrre molte planule nuove ed ancora
non avere rilevanti inincroci a causa d'eventi d'incroci durante
meiosi. Ogni polipo in una colonia subisce incroci separati
nella produzione di gameti, e così una colonia di alcune
migliaia, o anche milioni di polipi può produrre teoreticamente
la metà del numero di nuove colonie vitali, geneticamente
diverse, da solo.
 |
Questa caratteristica ha molte importanti
implicazioni. Prima, ci fa capire come coralli possono essere
efficaci in termini di tenere le barriere popolate, e questo
e certamente la ragione per loro successo eterno. Secondo,
è testimonianza di vantaggi dell'ermafroditismo e del
modo di vivere in colonie. Terzo, dovrebbe essere apparente
come vantaggioso potesse essere di riuscire produrre tutti
i coralli richiesti per il mercato acquariofilo, con la riproduzione
sessuale e loro allevamento. Letteralmente, uno o due colonie
di una specie potrebbero provvedere per ogni corallo richiesto
dal mercato, per sempre. In altre parole, raccolta di un migliaio
di colonie coralline di specie diversa potrebbe essere l'unico
impatto distruttivo, da parte di mercato acquariofilo, sulle
barriere da ora in poi. Questa quantità riuscirebbe
a soddisfare la stessa diversità di specie disponibile,
e continuamente. Al contrario, quest'anno un milione di colonie
coralline sarà raccolto dalle barriere per soddisfare
solo il mercato di Stati Uniti. La maggior parte di loro non
sopravvivrà, e dei superstiti la maggior parte non
si riprodurranno mai.
La maggiore differenza nella storia di
vita di coralli non è tra gonochorismo ed ermafroditismo,
ma nella differenza tra "spawners" (fertilizzazione
esterna) e "brooders" (fertilizzazione interna).
Queste differenze includono il trasferimento di zooxantelle
alle larve, tempi di dispersione larvale, modi di dispersione,
la variabilità genetica, percentuali di diverse specie
e l'evoluzione.
"Brooders" producono planule
(larva pelagica) che nel momento di rilascio, sono immediatamente
in grado di stabilirsi e fare metamorfosi in corallo giovanile.
Loro sono generalmente più grandi delle larve prodotte
da non-brooders, e contengono una "cultura di partenza"
di zooxantelle. Come tali, loro tendono a non disperdersi
per lungo tempo o molto lontano. Di solito si stabiliscono
e fanno metamorfosi vicino al genitore o all'interno della
stessa comunità locale. È interessante che,
anche se queste planule sono potenzialmente capaci di disperdersi
più lontano a causa delle zooxantelle che li forniscono
con energia, loro generalmente non fanno cosi. Non tutti gli
"brooders" sviluppano le planule all'interno della
cavità gastrica del polipo. Alcune specie sviluppano
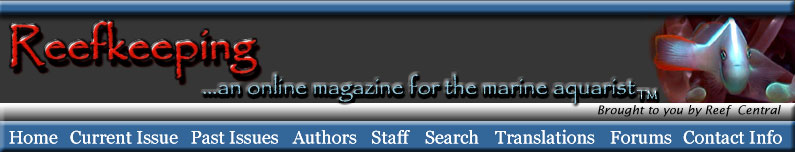
le planule sulla superficie della colonia, sotto il muco.
Coralli molli, come Clavularia e Briareum prendono
parte di queste specie.
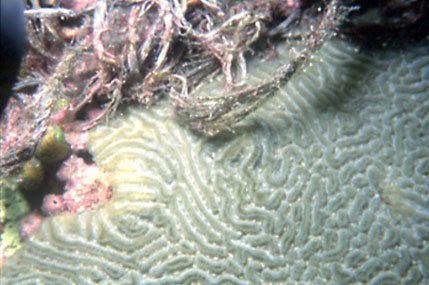 |
"Spawners", al contrario si fertilizzano
esternamente, e le planule si sviluppano nella colonna d'acqua.
L'intervallo che passa prima che loro sono in grado di stabilirsi
dipende dalla specie, e può dipendere anche da aspetti
ambientali. Questo tempo può variare da ore a settimane.
Perché sperma ed uovo sono separati e si fertilizzano
esterno al polipo del genitore, zooxantelle devono essere
acquisite dalla colonna d'acqua. Dipendendo dalla durata dell'acquisizione
di zooxantelle, e le caratteristiche specifiche di planule
della certa specie, la fase pelagica prima di essere in grado
di stabilirsi può durare da meno di un giorno a più
di 120 giorni. Coralli "broadcast spawners" tendono
a stabilirsi e fare metamorfosi lontano dalle comunità
locali, ma si pensa che di solito si stabilisca, non più
lontano che le comunità vicine (con ovvio potenziale
d'andare più lontano). In ogni modo, quest'aspetto
è raramente stato tentato di essere misurato, perché
è molto difficile seguire gli spostamenti della planula
individuale nello stato pelagico.
La Magia d'Essere una "Sincrona, Simultanea,
Ermafrodita, Esternamente Fertilizzata, Coloniale Specie Corallina"
Un bel boccone della verbosità!
Come sarà discusso più dettagliatamente in seguito,
"broadcast spawners" spesso rilasciano gameti en
masse, anche noto come riproduzione sincrona di massa
ovvero, fra un periodo che varia da ore a giorni, molte specie
liberano i loro gameti contemporaneamente. Questo è
chiamato"swamping" (inondazione), e si pensa che
massimizzi il numero delle fertilizzazioni riuscite, e sta
riducendo il numero di uova che predatori potrebbero consumare
sovraccaricando le loro abilità di consumarli tutti.
La maggior parte d'ermafroditi corallini simultanei non rilascia
separatamente uova e sperma, ma li rilascia in pacchetti d'uovo-sperma.
Alcuni contengono diverse centinaia d'uova che sono circondate
da un pacchetto di sperma. Dopo un certo periodo che varia
da minuti ad ore, e dipendendo da aspetti ambientali, qualche
volta, i pacchetti si rompono e la fertilizzazione può
avvenire. Questo è importante, perché uova hanno
un galleggiamento corretto e andranno in superficie. Loro
periodo di capacità fertilizzante e abbastanza lungo
in acqua marina. Sperma, in ogni modo, è diffuso rapidamente
in acqua, non è capace di galleggiare, e ha un periodo
di capacità fertilizzante corto in acqua marina. Se
uova e sperma non sono rilasciati precisamente nello stesso
momento, e se la fertilizzazione non avvenisse nell'immediato
vicinato della colonia, accadrebbe probabilmente, una percentuale
di successo di fertilizzazione più bassa. Come uova
e pacchetti d'uovo-sperma galleggiano, loro formano macchie
sulla superficie d'acqua che può andare alla deriva
per giorni, allontanandosi cosi molti chilometri dal luogo
di rilascio.Galleggiando cosi insieme, dopo la rottura dei
pacchetti d'uovo-sperma, la probabilità che succede
la fertilizzazione e più grande. E molto più
grande anche la probabilità di mescolamento genetico.
Inoltre, come planule di "broadcast spawners" possono
disperdersi più lontano, loro tendono a persistere
di più durante l'evoluzione. Questo probabilmente incide
poiché la maggior parte di coralli sono "broadcast
spawners", sincroni, ermafroditi simultanei. La percentuale
di successo per la fertilizzazione e mantenimento genetico
della specie e metapopolazione è il più alto.
Sincronismo
Molta attenzione è stata data ai
fattori che danno luogo alle quasi mistiche riproduzioni sincrone
di massa, che accadono sulle barriere coralline in tutto il
mondo. Prima di continuare vorrei fare una piccola prefazione
dicendo alcune cose: Prima, non tutti i coralli, né
tutte le barriere coralline, abbiano eventi di riproduzioni
di massa; Secondo, non tutti i coralli né tutte le
barriere coralline, depongono le uova contemporaneamente;
Terzo, non tutti i coralli o barriere coralline, sembra di
dipendere dagli stessi fattori che generano la riproduzione
di massa. La prima riproduzione di massa non fu scoperta ufficialmente
fino ai primi 1980! Da allora in poi, e con sempre di più
subacquei e ricercatori nelle acque, sta diventando molto
chiaro che tanti, se non tutti coralli sulle barriere partecipano
in questi eventi sorprendenti. Per chiunque interessato, il
video, Coral Sea Dreaming, un lungometraggio sulla
riproduzione di massa alla Grande Barriera Corallina in Australia,
che include coralli, stelle marine, oloturie, e policheti
in una mostra subacquea che è troppo bella perché
possa essere descritta.
Come ho gia scritto nella prefazione sopra,
e al dispiacere di tanti acquariofili, fattori esatti che
provocano la riproduzione di massa non sono ancora capiti
pienamente. Molti acquariofili credono erroneamente che loro
provocheranno la riproduzione nei loro acquari mettendo sopra
una luce blu che imita "fasi lunari". Sfortunatamente,
io temo che questo non farà molto. Quello che segue
e un sommario di quello che sembra essere la sequenza generale
d'eventi necessari per i coralli, a maturare e rilasciare
i loro gameti.
 |
Coralli, come gli altri organismi impiegano
tempo per giungere alla maturità riproduttiva. Diversamente
da creature umane, non esiste periodo fisso prima e durante
la "pubertà". Per alcune specie, età
sembra essere la causa importante; per esempio, la specie
di Xenia è stata trovata d'essere matura per
la riproduzione approssimativamente dopo un anno di vita,
mentre colonie femmine di Sarcophyton spp. arrivano
a dieci, dodici anni prima di essere mature.. Per altre specie,
sembra che una taglia fissa deve essere raggiunta prima la
colonia diviene matura per la riproduzione. Per altri coralli,
loro devono avere un'area di superficie che eccede una certa
taglia critica, mentre per altri può essere lunghezza
di ramo. Per ancora l'altra specie, appare che la densità
di polipo può essere la causa decisiva per la maturità
sessuale; ovvero, una volta un certo numero di polipi è
stato formato, la colonia può cominciare produrre gameti
senza riguardo all'area di superficie della colonia o la lunghezza
dei rami - processi che sono confusi da aspetti della crescita
come la velocità di calcificazione, la forma di crescita,
e la densità scheletrica.
E importante capire che, anche se una colonia
può essere di un'età o taglia corretta, o la
densità di polipo raggiunta, non c'è nessuna
garanzia che la colonia è, o diverrà feconda
(capace di riprodursi). Produzione di gameti richiede una
gran quantità d'energia, ed un corallo "broadcast
spawner" può rilasciare 25-75% della sua biomassa
deponendo le uova. Così, produzione di gameti richiede
un'eccedenza significativa d'energia oltre quella richiesta
per metabolismo e la crescita. Durante gametogenesi (produzione
dei gameti), è stato trovato, che molti coralli possono
anche fermarsi ad alimentarsi, così che eccesso deve
essere acquisito prima dell'inizio della formazione di gameti,
ed e acquisito da fotosintesi in coralli zooxantellati. In
ogni modo, gameti sono ricchi di proteine e così fotosintesi
può essere più importante per mantenere semplicemente
il corallo durante la metà dell'anno mentre sta producendo
gonadi mature, piuttosto che come un gran contributo alle
gonadi direttamente. Alcuno stress, danno, o la mortalità
parziale della colonia può consumare abbastanza energia,
tale che la produzione di gameti si è arrestata e gameti
sono stati riassorbiti, o può lasciare la colonia ad
una taglia o la densità di polipo, sotto il minimo
richiesto per riproduzione. In dei casi, una colonia può
essere in parte feconda, con le grandi aree sane espansive
della colonia che rilascia gameti, e con aree meno sane che
non hanno polipi fecondi.
Si pensa che temperatura ha il ruolo importante
nella segnalazione per gonadi e produzione di gameti. Generalmente,
come temperature cominciano a salire, gonadi cominciano a
maturare. Questo processo può durare la metà
dell'anno o più, ed aree con temperature annuali variabili
o molto stabile presenta piuttosto un mistero. È anche
noto che l'inverno è un periodo della crescita ridotta,
ed inizio di primavera spesso un periodo dell'abbondanza di
plancton. Forse coralli usano questo periodo, per ammassare
il cibo ricco d'azoto richiesto per il periodo di produzione
di gonade che accade durante l'estate. Questo processo di
maturazione è tipico per la maggior parte di coralli
sulla barriera, in che la maggioranza sembra d'avere eventi
di riproduzioni di massa annuali. Molte barriere e coralli
possono deporre le uova biennalmente, o semestralmente, ed
alcuni coralli possono deporre le uova anche su un ciclo mensile.
Questo chiaramente indica che si deve imparare di più
sul comportamento riproduttivo dei coralli. D'altra parte,
anni anomali con riguardo alla temperatura, danno spesso luogo
agli eventi di deposizione delle uova fuori dell'aspettato
periodo, o loro non accadono del tutto quell'anno. Questo
ci dà la credenza supplementare al ruolo di temperatura
d'acqua in maturazione sessuale dei coralli di barriera.
 |
Come temperature d'acqua giungono ai loro
massimi annuali, i gameti sono prodotti dalle ovaie e gonadi,
ora maturi, e risiedono all'interno dei coralli finché
un evento "di scatto" non provoca la loro liberazione.
Come indicato sopra, la deposizione delle uova tipicamente
succede nel mese di, o il mese dopo, la più calda media
temperatura mensile d'acqua, nelle riproduzioni di massa annuali.
In ogni caso, la "fama" d'eventi delle riproduzioni
di massa sono legate alla periodicità lunare. Si pensa
che la fase lunare offre le indicazioni per il tempismo di
liberazione massiccia di gameti. In altre parole, durante
il mese della temperatura d'acqua più alta, il giorno
e l'ora della liberazione dipende dalla fase della luna. Si
pensò, basato su primi rapporti, dai luoghi limitati
delle riproduzioni massicce, cha la riproduzione accadde durante
la notte o due ore dopo la luna piena. In ogni modo, con più
rapporti e le documentazioni, deporre le uova può essere
veramente correlato a fase lunare, ma varia significativamente
attraverso ubicazioni.Ha più senso, che liberazione
dovrebbe avvenire quando la luna è nuova e le acque
sono più scure per minimizzare perdita d'uova dalla
predazione. In ogni caso, io non sono consapevole che alcun
vero modello, ed esatto, ed emerso, per fase lunare, che incorpora
la vera maggioranza di barriere. Sfortunatamente per acquariofili,
cui i coralli, probabile vennero da molte barriere diverse,
l'uso di una luce che simula le fasi lunari, non avrà
probabilmente l'effetto di provocare la riproduzione massiccia
nella vasca, anche se la temperatura nella vasca e controllata
e simula cicli stagionali, ed anche se i coralli sono sessualmente
maturi e fecondi.
Inoltre, aspetti chimici sono coinvolti
nella deposizione delle uova e riproduzioni di massa, anche
se le indicazioni e fattori essati stiano cominciando ad essere
scoperti. Attiranti dello sperma sono stati isolati in un
numero di specie, e questi "segnali" trasmessi in
acqua possono essere importanti nel provocare la riproduzione
nell'altra specie. In somma, un numero d'altri fattori è
stato proposto, o è stato determinato che stia contribuendo
a, o essendo responsabile per, i vari aspetti del comportamento
riproduttivo. Un elenco dei tanti fattori coinvolti nella
riproduzione corallina di massa si trova nella Tabella 1.
|
Periodicità
annuale
|
Periodicità
stagionale
|
Periodicità
diurna
|
|
Periodicità
lunare
|
Segnalazione
chimica
|
Temperatura
d'acqua
|
|
Influenze
della marea
|
Differenze
latitudinali
|
Genotipo
di specie
|
|
Fattori
nutritivi
|
Disponibilità
della luce
|
Fasi
lunari
|
|
Grandezza
della colonia
|
Densità
di polipo
|
Età
della colonia
|
|
Salute
della colonia
|
Qualità
d'acqua
|
Salinità
|
|
Durata
del giorno
|
Struttura
della Comunità
|
Movimento
d'acqua
|
|
Campi
gravitazionali
|
Abbondanza
di predatore
|
Durata
della notte
|
|
Tabella
1. Fattori proposti o decisivi che contribuiscono
alla maturità sessuale dei coralli di barriera.
|
In particolare, un'area in quale credo
e merita più studio fu proposta come un risultato d'annuale
riproduzione massiccia e sincronizzata in coralli del Mar
Rosso dove la temperatura era veramente variabile durante
l'anno. Polipi corallini non sono capaci di percepire l'intensità
della luce, ma le zooxantelle sono capaci. Esiste in ogni
modo, la domanda considerabile sull'abilità di queste
alghe intracellulari di percepire cambi nella densità
di flusso dei fotoni sulla scala dell'illuminazione ai livelli
di luce lunare, anche su una luna piena. Zooxantelle risiedono
all'interno di tessuto corallino dello spessore e l'opacità
diversa, sotto uno strato di muco variabilmente spesso, che
contiene livelli variabili d'altri microbi fotosintetici e
non riceve nient'altro che una frazione della luce che arriva
alla superficie corallina. Profondità, torbidezza dell'acqua,
nubi, foschia, passaggi d'aerei Israeliani, o qualsiasi numero
d'eventi minori potrebbe cambiare i livelli dell'illuminazione
lunare, fino al punto che e quasi impossibile credere in possibilità
che zooxantelle o coralli riescono a percepire che in una
notte particolare la luna e piena o e una luna nuova. L'evento
di deposizione delle uova spesso accade, sebbene non sempre,
senza riguardo se la notte della deposizione e nuvolosa o
quanto l'acqua e torbida o profonda, o quanto una colonia
ombreggia l'altra. Un organismo per avere una risposta biologica
alla luce richiede un fotorecettore ed un'immediata e quantificabile
risposta per il suo sistema nervoso. In coralli gli unici
fotorecettori sono le zooxantelle, e la loro risposta, la
produzione di fotosinteticamente prodotte sostanze chimiche,
né è specifica abbastanza né c'e abbastanza
per tale uso.
 |
In ogni modo, e come lo studio menzionato
sopra lo suggerito, c'è un fattore che non varierà
annualmente, e livelli dell'illuminazione e periodi coinvolti
sarebbero percepibili da zooxantelle - lunghezza di giorno
o lunghezza di notte. Piante terrestri sono provocate a fiorire,
fruttare o cambiare colori di fogli in autunno, basato su,
se loro sono piante di giorno lungo (notte corta), giorno
intermedio, o giorno corto (notte lunga). In altre parole,
quando lunghezza di giorno o lunghezza di notte giunge ad
un valore critico, fitocromi provocano processi di traduzione
dei segnali che causano la produzione degli ormoni ed altri
segnali, che a turno causano processi comportamentali e biochimici.
Sento che la futura ricerca troverà che zooxantelle
sono responsabili per provocare la liberazione di gameti del
polipo corallino, producendo segnali chimici basati sulla
lunghezza del giorno o di notte. C'è ancora in ogni
caso, una domanda che rimane. Ci sono piante giorno-neutrali
che non rispondono a cambi di lunghezza del giorno o di notte,
e loro sono primariamente tropicali dove c'è poco o
nessuna variazione in lunghezza del giorno. Loro devono dipendere
da altri fattori che possono variare tra le specie. In ogni
modo, questo probabile spiega le variazioni nei tempismi delle
riproduzioni di massa su varie barriere, perché livelli
della luce lunare non sono stati trovati di avere una correlazione
forte attraverso le barriere, e forse più interessante,
perché le barriere equatoriali sembrano avere meno
prevedibili (o semi-annuali) eventi di riproduzioni massicce,
che quegli a latitudini più alte. Quest'idea a mio
avviso, non è intrapresa nella comunità di ricerca
corallina, ma merita investigazione.
Il Tempismo di Liberazione dei Gameti
 |
In generale, coralli rilasciano di notte
i loro gameti, anche se ci sono molti esempi di liberazione
di giorno ed alcuni, come Fungia sp. possono deporre
le uova giorno e notte. La maggior parte di riproduzioni di
massa si svolge in un periodo di minuti od ore, anche se liberazione
possa essere prolungata su molte notti consecutive. In casi
rari, come con Hydnophora exesa, la deposizione delle
uova può essere prolungata a settimane. Il tempismo
di riproduzione di solito si ripete, da anno ad anno; ed e
abbastanza preciso, che la riproduzione di specie coralline
può essere predetta quasi al minuto. Per esempio, la
Diploria depone le uova tra il 9: 00 ed il 10:00 di
sera e Montastraea depone le uova quindici, trenta
minuti più tardi, seguito un ora più tardi da
Stephanocoenia. Questa possibilità di prevedere
la riproduzione di almeno la specie corallina iniziale e una
cosa stupenda, anche se specie che la stanno seguendo forse
stanno reagendo ai segnali chimici.
Sistemazione e Trasformazione - Solamente l'Inizio
Sulla formazione riuscita di planule competenti,
fertilizzate internamente -coralli "brooders" o
nella colonna d'acqua - coralli "broadcast spawners",
esistono eventi che determinano se e dove le planule si stabiliranno
sulla barriera, e faranno metamorfosi in un polipo corallino
giovanile.Fino ad oggi, un numero di fattori è stato
proposto per spiegare gli aspetti ambientali richiesti per
dare l'inizio alla discesa della planula dalla colonna d'acqua,
per stabilirsi sul substrato. Fra i più studiati sono:
la rugosità (ruvidità) del substrato, profondità,
luce, biofilm prodotto da batteri e gli altri microrganismi
ed alghe coralline. Giorno d'oggi, si pensa, e ricerca suggerisce
che le alghe coralline sono notevolmente il più importante
fattore coinvolto. Specifici segnali chimici sono stati isolati
dalle alghe coralline che incitano sistemazione delle larve
di coralli e gli altri invertebrati, indicando cosi che la
profondità, luce, e biofilms sono secondari, se non
discutibili, nell'evento. Le variazioni riportate per gli
altri fattori sono spiegate facilmente in che varie alghe
coralline producono diversi segnali chimici, che variano nelle
profondità che loro occupano, abbiano diverso biofilm
associato, e possono crescere privilegiando substrati della
rugosità diversa.
 |
Una volta stabilite su alghe coralline,
planule tendono a stabilirsi in aree protette e strisciare
verso la luce. Il tempo per arrivare alla luce e diverso,
e probabilmente riflette il grado di competenza nel processo
di trasformazione che assicura la planula di recente stabilita,
che è in una posizione ottimale per sopravvivenza e
riflette la loro abilità cominciare a calcificare,
così come avere accesso a cibo, luce, e spazio. Specie
corallina, come Pocillopora damicornis produce larve
che possono rilasciarsi dal loro luogo di sistemazione, se
le condizioni non sono ottimali e ristabilirsi più
tardi con tempo (se substrato disponibile esiste!), indicando
che la sistemazione e processi di trasformazione sono dinamici
e non inflessibili. Dopo la sistemazione e trasformazione,
un piccolo polipo di corallo da solo affronta un periodo molto
incerto, con estremamente alte percentuali di mortalità.
In ogni caso, se riesce sopravivere, questo piccolo polipo
quasi irriconoscibile può, sul corso d'anni a secoli,
diventare una colonia di coralli capace di riseminare da sola
altre barriere coralline lontane migliaia di miglia - la storia
di vita straordinaria ed avventura di biologia.
Nel prossimo numero, la meraviglia della
natura descritta in quest'articolo sarà proseguita
nel descrivere anche gli eventi più straordinari…coralli
che depongono le uova negli acquari di barriera.
|