|
Vicious Viscosity
Many of the smaller
invertebrate or invertebrate-like organisms in reef aquaria
are well known to the aquarists maintaining those aquaria,
but many of the factors influencing these organisms are a
consequence of their small size and are difficult for aquarists
to comprehend. "Strange" things happen in water
when organisms are either very small or possess very little
mass. As I have mentioned many times in this column, as aquarists
we tend to interpret the world from our own perspective. There
is absolutely nothing wrong with this, but in doing so, we
must continually perform "reality checks" and realize
that our perspective may not always provide an appropriate
explanation of what is happening. In this particular case
potential misinterpretations are due to size. Humans are among
the largest animals on Earth; surely there are larger animals,
but just as certainly, well more than 99.9% of all animals
or animal-like organisms are smaller. And many of them are
VERY much smaller. This size difference does have some profound
consequences. Down to about the size of small aquarium fishes,
or perhaps small bristle worms, most of the things that we
take for granted about the world still work in ways that are
familiar. But for very small organisms, well… it is like
going through the looking glass after Alice: things can become
very bizarre.
With regard to aquarium organisms, what appears to be strange
is a result of the organisms' living in water. Water, like
air, is a fluid medium. The properties of fluids relative
to organisms depend upon the molecular properties of fluid's
constituents, and the sizes and shapes of the organisms. We
humans are used to air as one fluid medium with certain properties
and to water with another altogether different set of properties.
Yet, there are similarities between these media, and we can
move through both of them relatively easily. This ease of
movement depends largely on our mass. Once we start to move,
our mass ensures that we build up momentum, or inertia, which
allows us to move though the medium fairly easily.
When an organism's mass is low relative to the medium's viscosity,
however, movement may become very difficult, indeed. As an
example, if a human tries to walk in a wind blowing at about
100 miles per hour, his mass relative to the forces generated
by the air's movement becomes small, and it becomes quite
difficult for him to control his motion. Frictional effects,
particularly drag, increase and instead of being able to move
into the wind, he may find himself being blown along by it.
Water is a much more viscous medium than air, so similar viscosity
effects become apparent at much lower fluid velocities in
water than in air. A scuba diver who is not carrying any excess
equipment can swim against a water current flowing at between
1 and 2 miles (about 1.6 to 3.2 km) per hour without much
difficulty. On the other hand, if the diver is carrying a
lot of equipment, such as cameras and research gear, his effective
frontal area increases, and so does the water's resistance
to something moving through it. This resistance increases
with the square of the frontal area, so doubling the frontal
area increases the resistance by a factor of four. This means
that for a scuba diver carrying bulky equipment, swimming
even just enough to maintain a steady and stable position
in water moving at 1 mile (1.6 km) per hour may be difficult.
Relatively large scale oceanic water movements generated by
tides or winds can travel at velocities well in excess of
5 miles (8 km) per hour, and at these velocities a diver becomes
simply another piece of debris blowing along in the currents.
Allow me to share with you one of the shorter, but exciting,
moments of my research career. One of the research
sites that I used for my doctoral dissertation was the lower
intertidal region of beach on the eastern side of the channel
called "Point Washington Narrows" near Bremerton,
Washington. Tidal currents in this channel exceed 10 miles
(16 km) per hour. I once got a wonderfully wild and crazy
(i.e., very stupid) idea to see what was happening in my study
area while it was submerged during maximum flood tide. I strung
lines across the bottom, and thought I would be able to 1)
orient myself using the lines, which were labeled; and 2)
hold on and make observations of the animals, some venomous
snails I was studying. I did a lot of my research diving solo,
and this time was no different so, fortunately, I didn't have
to worry about a buddy. The water current in the area dissipates
at either end of the narrows, a channel about a mile (1.6
km) long, so that even if the current became too extreme,
I figured I could ride it out and be in relatively safe waters.
I waited for the current to reach its maximum, donned my dry
suit and entered the water. The results were ludicrous. I
was flushed so rapidly through the area that I never even
had a chance to grab onto the ropes I had strung over the
bottom. Additionally, visibility in that water was relatively
poor. By the time I could see the lines, I was past them and
couldn't grab them. In retrospect, this was probably a very
good thing. Had I been able to grab one and hang on, I would
have been flapping like a torn flag in a gale. I did see my
study site… it was about 530 feet (160 m) long. I passed
over the entire site in less than six minutes, and believe
me, in the murky, cold water of that December day in 1974,
that was a real cheap thrill! However, it did give me a really
good appreciation for the problems of being small in a viscous
medium. Ah, to be young and dumb(er) again…
The point of this recollection is not just to illustrate
the idiocy of youth, but to note that most marine organisms
experience similar effects at much lower velocities. This
is because they have a much lower mass so, relative to their
sizes, water is much more viscous. This viscosity is due primarily
to the dielectric properties of water molecules, which tend
to cause water molecules to attract other water molecules.
In effect, water molecules "stick" to one another.
This "stickiness" of water has a great deal to do
with both the movement of small organisms in it and its movement
through small channels such as the interstitial pores of a
deep sand bed, or the spaces inside an animal where it is
moving as blood. Water's properties have a great deal to do
with the design and functionality of organisms, both on the
outside and on the inside.
Movement
|
Figure 1. Pseudopodia are generally too small
to see with the unaided eye. However, the sessile red
foraminiferan Homotrema rubrum, which is common
in aquaria, catches particulate organic material on
pseudopodia that it extends out into the water. These
are visible as fine, threadlike extensions at the ends
of the foram's branches.
|
|
As far as organisms are concerned,
probably the most basic functional property that viscosity
affects is water movement, and their small size combined with
water's odd properties go a long way toward explaining some
of the basic biological attributes of tiny organisms. The
most fundamental type of movement found in tiny animal-like
organisms is done by extending their body's entire surface.
This type of movement is seen to best advantage in organisms
such as amoebae. Small organisms, on the size scale of amoebae,
are generally less than about 0.1 mm (1/250 inch) in diameter.
Often these organisms are not comprised of multiple cells,
and because of this they are not animals, which by definition
are multicellular. Such organisms belong to several different
and unrelated lineages, but we often lump them together by
calling them protozoans, a name that means "first animals,"
as at one time they were considered to be simple animals,
possibly similar to the ancestors of "true" animals.
Protozoans tend to have a lot of peculiar bodily structures
derived from the single cell that constitutes their body but,
of course, they lack things such as muscles and skin and other
cellular derivatives. All amoebae and their relatives; organisms
such as true free-living Amoeba;
many similar parasitic forms such as Entamoeba
histolytica, which can cause dysentery in humans;
and the foraminiferans
common in marine environments and aquaria can move by extending
blobby, hair-like, or even net-like extensions of their body's
surface called "pseudopods,"
a name meaning "false feet." Pseudopods typically
move because of some interesting properties of the organism's
structure. The outer part, called the "ectoplasm"
of the organism, just inside the exterior surface's covering,
or cell membrane, is altered chemically and becomes more fluid.
Internally, the fluid that constitutes the internal bodily
"goo," or cytoplasm, is filled with all sorts of
odd chemicals, including a network of long and interconnected
proteins connecting all the various parts of these tiny organisms'
internal structure. These proteins are altered, causing some
of them to contract, which forces the cytoplasm out into the
more fluid regions. This, in turn, forces the cell membrane
outward into a long extension, "the pseudopod."
Pseudopods are quite effective locomotory devices and are
surprisingly mobile and agile. In laboratory exercises, students
are often quite amazed to watch what appears to be a "sluggish"
amoeba capture and eat far more rapidly moving protozoans
or even small animals such as rotifers. It is apparent from
watching the movement of pseudopods that they don't move through
the water easily. It provides a lot of resistance, and instead
of rapidly flowing outward, they ooze slowly through it, often
taking seconds to move just a few micrometers. Additionally,
water's viscosity can be seen to influence the locomotion
of amoeboid organisms in another way. If the water is relatively
still, amoebae may be seen moving vertically through the water
column. They are not swimming as much as simply climbing up
into the viscous water. To these organisms water is about
as fluid as thick corn syrup is to us, and they can quite
easily climb up into it.
Pseudopodial movement is found in most free-living unicellular
organisms, with the exception of diatoms. It is found also
within the bodies of most multicellular organisms because
these organisms contain cells that move around freely within
and through tissues. For example, corals and other cnidarians
contain "interstitial cells." These are cells that
are termed to be "undifferentiated," which simply
means that they don't look like anything other than a generalized
animal cell. In other words, they look like an amoeba. They
act like one, too. They move from place to place within the
coral and can perform all sorts of tasks, depending on the
animal's need. They can help repair injuries or turn into
an egg, or turn into any other type of specialized cell. Aquarists
also have many cells in their bodies that move by the use
of pseudopods. Probably the most abundant of these are white
blood cells, especially macrophages,
which look and act quite like amoebae.
Cilia
Although pseudopodial movement is
ubiquitous, probably the most widespread and common locomotory
"device" used by organisms is a microscopic beating
filamentous structure referred to, variously, as an undulopodium,
a cilium or a flagellum. The main structural difference between
a cilium and flagellum is length; flagella are typically many
times longer than cilia. The mouthful term, undulopodium,
used in a few texts and references, is a term devised to encompass
both cilia and flagella. These structures are microscopic
and therefore invisible to the unaided eye. In fact, they
are so small that their basic internal structural attributes
are invisible even with the best light microscope. As a consequence
of this, an understanding of ciliary structure and function
had to await the development of the electron microscope, which
can magnify structures to a much greater degree than can a
light microscope. Although cilia and flagella have been known
for several hundred years, their main structural components
and mode of operation remained undiscovered until the latter
half of the twentieth century.
A cilium or flagellum is an extension of a cell's surface.
Although hair-like in proportion, it is not a hair in the
sense of a mammalian hair. Mammalian hairs are non-living
proteinaceous excretions of glands located in the skin, and
are thousands of times larger than either cilia or flagella.
The cellular surface extension that comprises the cilium or
flagellum is very narrow and roughly circular in cross-section.
These structures are typically many hundreds of times longer
than wide. As they are extensions of the cellular surface,
they are covered externally with a cell membrane continuous
with the membrane covering the rest of the cell. Internally,
both structures possess a rather consistent anatomy. They
are comprised of nine tiny tubular molecules, called "microtubules,"
which are arranged in a helical pattern. In the center of
this helix of nine microtubules are two more microtubules.
This rather peculiar arrangement of 9+2
microtubules is remarkably consistent throughout organisms
possessing cilia or flagella. The internal
microtubular arrangement is the same in the flagellum
of a green unicellular microalga such as Euglena
(1),
or in a dinoflagellate,
or in a human sperm cell, or in an epithelial cell of a coral.
This unity of structure is considered to be one of the fundamental
unifying factors showing that all life is related.
|
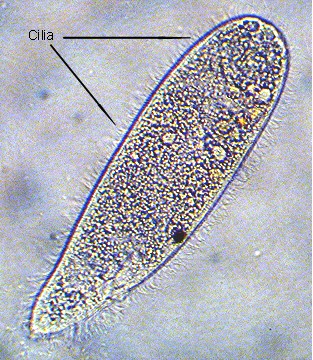
Figure 2. Microscopic ciliated protozoans, such
as this small organism, are common in aquaria. This
individual was about 0.05 mm (1/500 inch) long. The
cilia are visible covering its surface.
|
To explain that everything has its cost, we often use the
aphorism, "There is no free lunch." That truism
is as valid with ciliary locomotion as it is with the more
familiar muscular movement. It takes energy to make motion.
Cilia or flagella move by the application of chemical energy,
most often derived from the breakdown of an adenosine triphosphate
molecule (ATP) to an adenosine diphosphate (ADP) molecule
and a free phosphate ion. The
release of that phosphate ion liberates chemical energy that
is used to move the nine external microtubules relative to
each other and relative to the internal pair. That
movement causes the entire structure to bend and flex.
The bend and flexion is done in a specific "twisting"
pattern resulting in a relatively stiff "power-stroke"
phase and a rather flaccid "recovery-stroke." If
the power stroke is from a cilium, the water is generally
moved downstream toward the tip of the cilium or the animal
is moved in the opposite; remember Sir Isaac Newton's third
law regarding direction of forces. If the power stroke is
from a flagellum, the resultant thrust vector is much more
difficult to predict, and may be either direction along the
flagellum or even lateral to the axis of movement. During
the power stroke, the hair-like structure pushes against the
water molecules and acts to move either the water or the organism.
The only reasons a cilium or flagellum actually can function
are because of the water's cohesiveness and the small size
of the organism using it. When we view the movement of a cilium
or flagellum, it often appears as if a fine thread is moving
in fluid water; intuitively, we know that such a movement
would generate very little thrust. Actually, the ciliary or
flagellary action is more like an oar sculling through water
to push a boat and, on the scale of a microscopic organism
with very little inertia, the force generated by a cilium
or flagellum is quite sufficient for movement.
Nevertheless, the thrust developed from one flagellum or
cilium is relatively small, but so are the organisms they
move. On a relative scale of body lengths per second, many
flagellated organisms with a single flagellum can move rapidly
over relatively great distances and, of course, all of the
readers of this column are the result of just such a race
run by many small flagellated spermatozoa. While most organisms
tend to bear flagella in more-or-less small groups, generally
from 1 to 10 per microscopic organism, there are exceptions
to this, and some flagellated protozoans found in termite
and wood-roach guts may be covered in thousands of flagella
(See this diagram
and image
of Trichonympha, a flagellate found in the guts of
termites).
Cilia, on the other hand, are more typically found in huge
numbers. Many small mobile organisms such as ciliated
protozoans, the numerous free-living flatworms, acoelomorphs
(the so-called "acoel flatworms" such as the "oh,
so familiar" red planarian, Convolutriloba retrogemma
which is not even a typical flatworm, let alone a planarian),
and ribbon worms are completely covered with them. Additionally,
discrete bands of cilia are often found on larvae and are
quite capable of moving them rapidly and over great distances.
Small, ciliary swimming organisms are exceptionally common
in the world's oceans, and are often the foods of many suspension-feeding
organisms such as stony corals, soft corals or other filter
feeders.
|
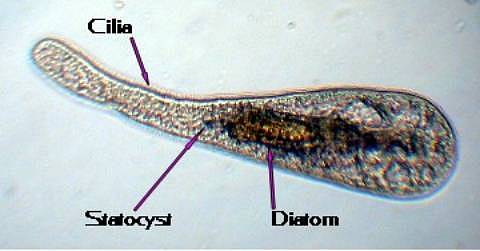
Figure 3. Acoelomorphs, animals which used to
be called "acoel flatworms," may reach a length
of 3 to 4 mm (1/8th to 1/6th inch), but most of them
are smaller. Although they have a cellular epidermis
covered in cilia used for locomotion, the smaller acoelomorphs
look a lot like ciliated protozoa. They can be easily
distinguished from ciliates, however, by the presence
of a spherical statocyst near their front end.
|
Ciliary locomotion on surfaces results in the characteristic
"gliding" movement typical of many small aquarium
animals, seen to best advantage on aquarium walls. In these
cases, the animal is moved by its cilia in a manner quite
like that of a microscopic ciliated protozoan. It may be a
bit surprising, but many larger animals also utilize cilia
for locomotion. Among animals living on the ocean's bottom,
it is likely that the most massive animals to move by ciliary
means are many large snails that move using a ciliated foot.
These snails move with a gliding motion created by the ciliated
epidermis covering their foot. In aquaria, the most common
snails to move in this manner are probably the meat-eating
scavengers in the genus Nassarius.
|
Figure 4. A specimen of Lewis' moon snail, Polinices
lewisii. These animals move exclusively by cilia
which cover their large expanded foot (very little other
than the foot is visible in this image; only a small
portion of the shell is visible at the top of the animal).
These moon snails are among the largest, if not the
largest, animals to move exclusively by ciliary means.
The animal is over 20 cm (8 in) long and weighs about
450 g (1 pound).
|
Most mobile and sessile marine animals that lack an exoskeleton
possess cilia on their epidermis. These cilia move water over
the epidermis and continually bathe the animal's surface in
clean sea water. Such an arrangement is found on animals as
varied as corals and sea stars and is probably the most effective
way of maintaining the cellular surface in good condition.
The water movement so generated acts to facilitate diffusion
of materials both into and out of cells. Such self-generated
water movement is particularly important to sessile animals
such as corals. In many normal marine environments, the water
flow near the substrate is almost nil due to the water's viscosity
and the development of a water layer called the benthic boundary
which gets created close to all surfaces in areas with smooth
or laminar water flow. Boundary layer effects may be seen
in air as well; the equivalent to the underwater benthic boundary
layer is found over the surface of a moving fan blade. Even
though the blade may be moving a lot of air, the air directly
over the blade's surface is stagnant, and dust, which can
easily be blown off the blade, will settle on it, even as
it is moving. Similarly, laminar water current flowing over
an organism will not move water in the benthic boundary layer
that surrounds the organism and without some means to generate
such currents, even on a small scale, organisms would suffer
problems in breathing and waste transfer.
Conclusion
An understanding of the basic cellular
means of locomotion, whether or not they move the organisms
or water around or even function within organisms, is fundamental
to the understanding of the "biology" of all organisms.
On the basest level cilia or flagella move small organisms.
They also move water around larger organisms, assisting in
respiration and waste removal. On top of this, they are also
found within many organisms moving fluids from one portion
of the body to another. For example, in echinoderms such as
sea stars and their relatives, cilia move the body fluids
around and act as the motive force for internal circulation.
Although these animals don't have a circulatory system with
a muscular heart, fluids move within circulatory channels
under the coordinated and cumulative action of millions of
beating cilia.
Ciliary and flagellar locomotion are the fundamental means
of rapid locomotion by small organisms, and were undoubtedly
the means used by the first small mobile animals, animals
apparently quite like acoelomorphs (1,
2,
3)
in size and structure. However, this type of locomotion is
limited by the size and, to some extent, the shape of the
organism. For animals to get big and to be successful, other
means of movement were necessary. Next month, I will discuss
locomotion by muscular means and how the development of discrete
muscular layers first allowed effective crawling and then
rapid swimming.
|
|
Figure 5. Although acoelomorphs look like
flatworms or ciliated protozoans, they are not closely
related to either group, but instead appear to be a
remnant of the ancestral stock of all bilateral animals.
This small, 0.05 mm (1/500 inch) long, acoelomorph has
eaten a diatom, which is visible in its body. All acoelomorphs
move using cilia.
|
|



)