|
Introduction
In the earlier part of this article,
I described the role of oxygen in seawater and its potential
effects on marine species. I noted, particularly, that hypoxia
is likely to occur in reef waters, within coral colonies,
at the coral tissue's surface, and that hypoxia potentially
affects species exposed to it. The scientific literature suggests
that such conditions might be relatively common at night when
respiration is high and primary oxygen production through
photosynthesis is not occurring. On coral reefs, although
oxygen levels are supersaturated near the surface during the
day in shallow waters, they are often reduced by 40-90 percent
or more at night. Primary factors affecting the oxygen content
of reef waters at night include surface conditions and water
mixing, as well as total community respiration. So how do
aquariums compare to reefs?
In this article, I report the results of numerous tests of
various water conditions in closed system aquaria. I utilized
various ways to "oxygenate" water and compared their
effectiveness. While the data I presented at the 2005 IMAC
conference were limited to some degree, I believe that enough
measurements have now been taken (including suggestions made
by attendees during the question and answer session following
my presentation of this material at the recent IMAC conference
in Chicago) to present a somewhat comprehensive picture of
oxygen dynamics in reef aquaria. The most notable additions
presented here are the effects of oxygen production in tanks
that are initially hypoxic. In the next article, I will continue
to report my results and discuss more natural oxygen dynamics
of several reef aquaria.
General Methods
I used a YSI Model 58 submersible
oxygen field probe to record oxygen levels in all tests. This
particular probe does not have a self-stirring BOD probe.
Without the self stirring device the probe must be exposed
to flowing water or be moved through the water at a rate fast
enough to obtain accurate measurements. More information on
this meter's specifications can be found here.
All readings were taken following the installation of a new
membrane, and the meter was calibrated by containment in a
100% humidity environment adjusted for Hg pressure at altitude,
according to instructions provided by the manufacturer. Another
calibration for a zero oxygen value was performed with a solution
of stirred water with a known volume of sodium thiosulfate,
an oxygen scavenger.
During the installation of the membrane, it is important
to avoid letting bubbles be trapped under the membrane or
readings will vary significantly. It is also important that
bubbles are not present in the water sample being recorded.
One very small bubble was present under the membrane, which
may account for some minimal variability in the results; although
after taking hundreds of readings and analyzing standard deviations
of the measurements, I can state that the readings are statistically
accurate and that any variance in replicated readings is insignificant
at a 95% confidence interval.
All readings were taken after a 10-minute stabilization period
for the probe in any given solution. In all measurements,
four readings were taken 15 seconds apart and the mean of
the values was recorded.
Experimental tanks included controls with sterile containers
and sterile seawater, either closed or open to the air, tanks
containing freshly mixed seawater using various water flow
or aeration devices but no organisms present and with or without
light, and various "reef aquaria" containing a variety
of organisms and apparatus. All tanks mentioned above contained
water at ambient levels of oxygen, either following mixing
in the case of the sterile controls, or those levels already
present in the tanks that were exposed to air or enclosed.
In addition, measurements were taken in tanks containing freshly
mixed seawater that had been made hypoxic by the rapid addition
of nitrogen gas to displace oxygen to a level below 20% of
saturation for all the conditions tested above.
All tanks contained seawater at 35psu as measured by a Reichert
salinity refractometer calibrated using double distilled water.
Although some variance in tank temperature occurred in the
tanks where lighting was a variable, the oxygen meter is temperature
compensating. The tank temperatures without lighting, and
some with lighting, were maintained at 26°C, with some
of my home systems reaching 26.6°C during the late afternoon.
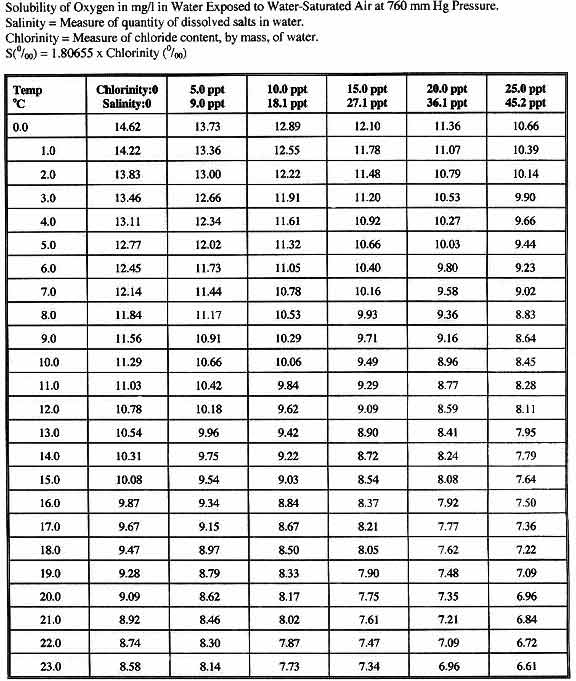
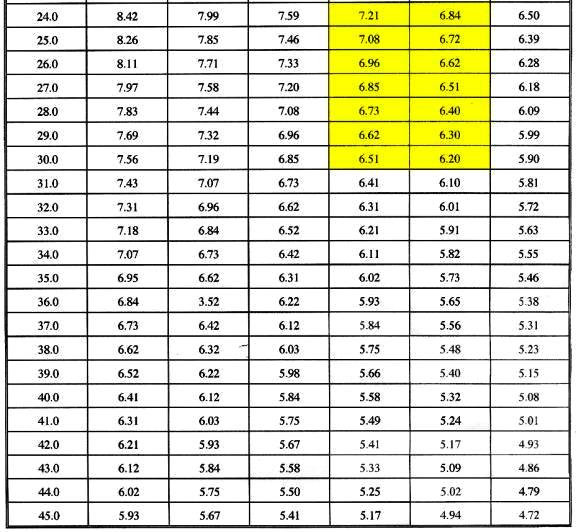
For informational purposes, oxygen saturation values at various
salinities and temperatures are provided in Table 1.
|
Table 1. Oxygen saturation values at various
temperatures and salinities. The yellow shaded regions
indicate the saturated oxygen values within the ranges
typically found in coral reefs and in reef aquaria.
All measurements in this paper are represented as a
percent of the oxygen saturation value at 35psu and
at a temperature of 26°C (approximately 6.66 mg
l-1).
|
Results
Control tank 1
Distilled water was autoclaved for
one hour prior to use. The salt source was an unopened 50-gallon
bag of Crystal Seas Bioassay Formula™ salt (Marine Enterprises,
Inc.). Seawater was prepared in clean two liter beakers, covered,
and stirred on a stir plate with a stir bar until completely
dissolved. Six liters of seawater at 35psu were filtered to
0.1µ through graded Whatman papers and then through
Millepore filters under a vacuum. Filtered seawater (1500ml)
was poured into a desiccation bowl that was triple rinsed,
then fitted with a stir bar and rubber stopper through which
the oxygen probe was placed into the seawater solution. The
bowl was placed onto a stir plate and its water was stirred
at a minimal rate in excess of that required for probe measurements
for a period of 240 minutes under ambient room light or in
darkness by covering the desiccation container with a thick
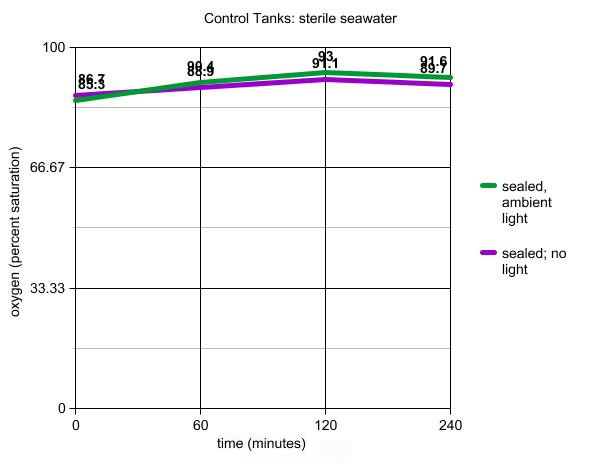
cardboard box. Results are shown in Table 2.
|
Table 2. Oxygen levels of sterile seawater placed
in sealed containers and either exposed to ambient room
light, or placed in darkness, for 240 minutes. Note
that the time scale is not equivalent between the 0
to 120 minutes and the 120 to 240 minutes segments -
measurements were taken less often during the third
and fourth hours due to their smaller rate of change
in oxygen level.
|
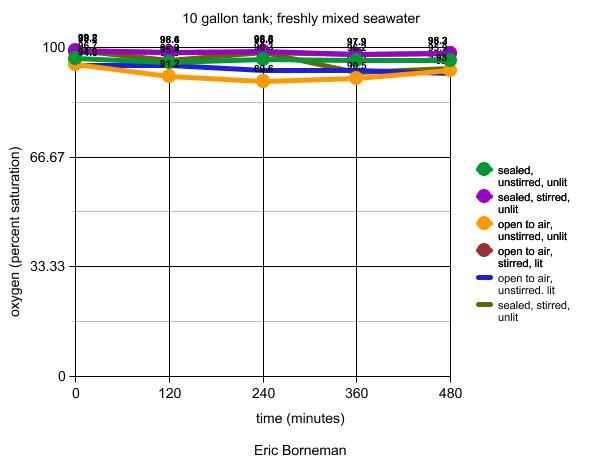
Experimental Tank 1
Ten gallons of seawater were mixed to 35psu using an unopened
bag of Instant Ocean™ salt and distilled water poured
into an acid-rinsed and neutralized 10-gallon glass aquarium.
As a first variable, the tank was either sealed with a sheet
of acrylic using removable tack adhesive to prevent air from
entering the aquarium, or it remained open to the air. A second
variable was the presence or absence of water circulation
by a single MaxiJet 1200™ (Aquarium Systems) placed at
the center of the tank's short end so that the flow moved
along the aquarium's long centerline. It was also placed just
far enough below the surface so that no air was pulled into
the pump. Visual inspection of circulation indicated a highly
turbulent flow. The oxygen meter was placed on the aquarium's
opposite end through a hole cut in the acrylic and sealed
with parafilm to fill any gaps. For stagnant tests the meter
was carefully rotated in a circle within the tank at a rate
allowing readings to be recorded; the parafilm provided a
flexible but airtight barrier to allow this movement. A third
variable was light. Lighting was provided by a single 18"
fluorescent fixture, or the room lights were turned off. While
not completely dark, there was too little light in the unlit
room to see the oxygen meter reading without a flashlight,
and it was assumed that this latter condition sufficed as
a "dark" condition. Results are shown in Table 3.
|
Table 3. Effects of water flow, light and free
air exposure on oxygen levels in seawater expressed
as a percentage of oxygen saturation at 35psu.
|
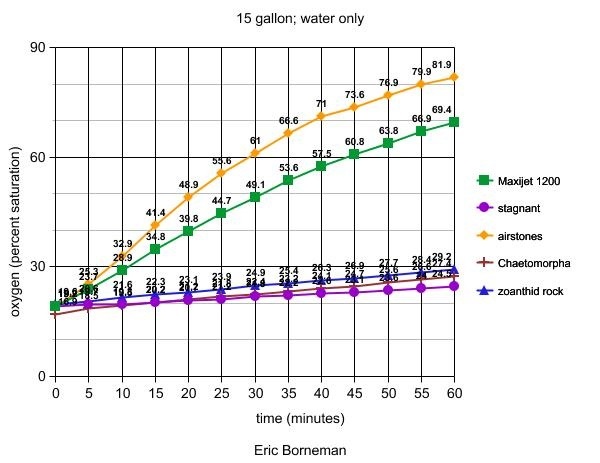
Experimental Tank 2
A 15-gallon (8" x 24" x 12") aquarium was
filled with seawater at 35psu prepared with deionized water
and Instant Ocean™ salt. Hypoxic water to below 20% saturation
was achieved by bubbling nitrogen gas into a powerhead's intake
to rapidly displace oxygen by saturating the tank's water
with fine nitrogen bubbles. A ten minute wait then allowed
all residual bubbles to rise to the surface and break so that
no bubbles would affect test readings by the probe. Results
are shown in Table 4. The conditions tested were as follows:
1. A single MaxiJet1200™ was placed at one
end of the tank, creating highly turbulent flow throughout
the water volume and visibly stirring the surface.
2. The tank was left open to the air without any
water flow and the oxygen probe was slowly moved at a rate
just fast enough to take a reading.
3. A single ceramic airstone was added at one end
of the tank using a Reno 400 air pump. Air bubbles were
coarse and did not reach the other end of the tank. The
power head was turned on for 15 seconds to mix the water
and a reading was taken by slowly moving the probe through
the water where no bubbles would affect the reading.
4. Thirty-seven grams of live Chaetomorpha
sp. algae were added to the tank and illuminated by two
15" 18-watt fluorescent daylight bulbs placed close
to the water's surface. The powerhead was turned on for
15 seconds to mix the water and a reading was taken by slowly
moving the probe through the water.
5. A four-pound piece of live rock approximately
40% covered with Protopalythoa sp. zoanthids and
60% with coralline algae was placed into the tank and illuminated
by two 15" 18-watt fluorescent bulbs placed close to
the water's surface. The power head was turned on for 15
seconds to mix the water and a reading was taken by slowly
moving the probe through the water.
|
Table 4. Effects of photosynthesis or aeration
devices on oxygen levels in hypoxic seawater (initial
O2 at 89.2% saturation) expressed as a percentage of
oxygen saturation at 35psu. When the airstone was placed
under the powerhead to mix the air and water into a
froth which was then allowed to settle before a reading
was taken, oxygen saturation reached 91.1% within 20
minutes.
|
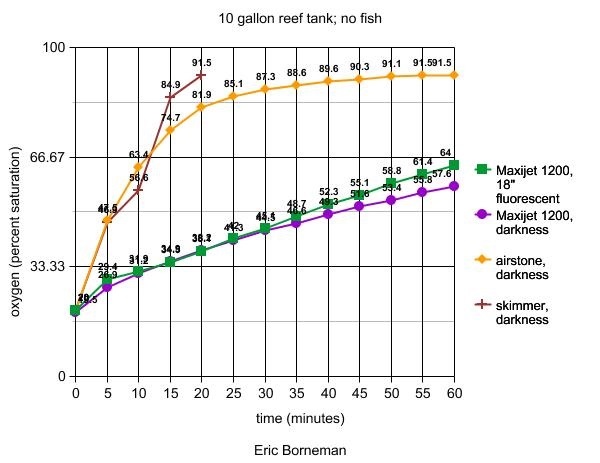
Experimental Tank 3
A 10-gallon reef aquarium established for three years in
my lab is described as follows:
Substrates:
5 cm medium grain aragonite, 3 kg live rock
Fish: none
Corals: small colonies:
Ricordea florida, Millepora sp., Porites
cylindrica, Zoanthus sp.
Rhodactis
inchoata
medium
colonies: Porites cylindrica, Protopalythoa
sp.
Invertebrates: five Trochus
sp. snails
Water flow: a single MaxiJet
1200™ powerhead (Aquarium Systems, Inc.)
Other filtration: CPR
BakPak™ skimmer
Water changes: none
Additions: kalkwasser
as replacement water
Lighting: a single 18"
fluorescent bulb (Lights of America, Inc.)
Tank 3
The oxygen probe was either stabilized with a ring stand
so that it remained submerged in the powerhead's water flow
(stirred conditions), or the meter was carefully rotated in
a circle within the tank at a rate that would allow for readings
to be taken. In the first set of experiments, nitrogen gas
was bubbled into the powerhead's intake to rapidly drop oxygen
levels until they reached approximately 20% saturation. Results
are shown in Table 5. The conditions tested were as follows:
1. A single MaxiJet 1200™ placed in the center
of the tank's short side and adjusted to create a lengthwise
circular water flow in the tank. The tank was illuminated
by a single 18" fluorescent daylight bulb (Lights of
America, Inc.).
2. A single MaxiJet 1200™ placed in the center
of the tank's short side and adjusted to create a lengthwise
circular water flow in the tank. All lights, including ambient
room lights, were turned off to simulate darkness.
3. A medium pore ceramic airstone connected to an
air pump (Supra™, Tetra Inc.) was placed in the center
of the tank. The powerhead was turned off. All lights, including
ambient room lights, were turned off to simulate darkness.
The meter was carefully rotated in a circle within the tank
away from any bubbles at a rate that would allow for readings
to be recorded.
4. A skimmer was placed on the tank (Remora hang-on™,
Aqua C). No powerheads or airstones were in operation. All
lights, including ambient room lights, were turned off to
simulate darkness. The meter was carefully rotated in a
circle within the tank away from any bubbles at a rate that
would allow for readings to be recorded.
|
Table 5. Effects of photosynthesis or aeration
devices on oxygen levels in a 10-gallon reef tank with
seawater made hypoxic (initial O2 at 90.8% saturation)
expressed as a percentage of oxygen saturation at 35psu.
|
Experimental Tanks 3 and 4
In addition to the tank described above, a second 10-gallon
reef aquarium established for three years in my lab is described
as follows:
Substrates:
5 cm medium grain aragonite, 3 kg live rock
Fish: none
Corals: small colonies:
Capnella sp. and Millepora sp.
medium
colonies: six Rhodactis inchoata
medium
to large colony: Zoanthus sociatus
Invertebrates: five Trochus
sp. snails
Water flow: a single MaxiJet
1200™ powerhead (Aquarium Systems, Inc.)
Other filtration: none
Water changes: none
Additions: kalkwasser
as replacement water
Lighting: a single 18"
fluorescent bulb (Lights of America, Inc.)
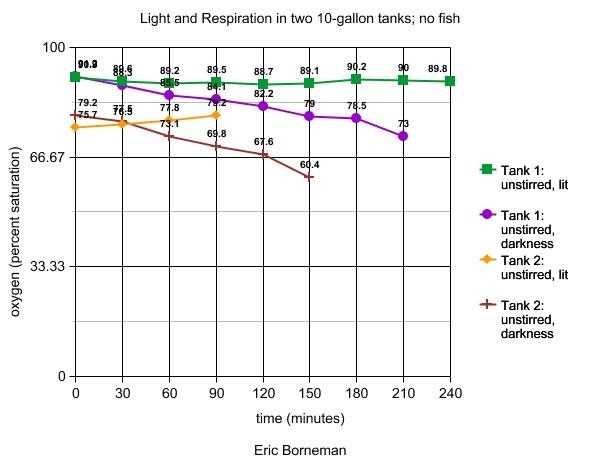
Tank 4
In this series of tests, the relative effects of photosynthesis
and respiration on the oxygen content of two 10-gallon reef
tanks were measured in the absence of any water flow or other
apparatus. Lights above the aquarium were either turned on,
or all lights, including ambient room lights, were turned
off to create darkness. Prior to beginning the experiment,
all tanks were operating with all apparatus as described above.
Results are shown in Table 6.
|
Table 6. The effects of photosynthesis and respiration
on oxygen levels in two 10-gallon reef tanks. Note that
tank 1, normally employing a protein skimmer, has an
initially higher oxygen level than tank 2, which is
run without a skimmer.
|
Experimental Tanks 5, 6 and 7
The following tanks are part of an interconnected six-tank
propagation system. It has been established for three months
and is described as follows:
Total system gallons:
600
Water flow: Ampmaster
2600™ centrifugal pump (Dolphin Aquarium and Pet Products,
Inc.) fitted with eductors at three of the six outflows
(one per tank); some tanks also have high volume powerheads
for circulation (6060 Stream, Tunze, Inc.; Seio 1100; Rio
pumps, Inc.)
Other filtration: MR-2™
protein skimmer (My Reef Creations, Inc.) powered by a Gen-X
MAK 4™ centrifugal pump (Pacific Imports)
Water changes: none
Additions: kalkwasser
as replacement water, calcium chloride, sodium carbonate,
sodium bicarbonate
Lighting: various, depending
on the species being cultured.
Tank 5
Size:
30-gallon
Substrates:
7.5 cm medium grain aragonite, small amount of live
rock rubble (<0.5 kg)
Fish:
none
Corals:
small colonies: 63 Capnella sp.
medium
colonies: three Capnella sp.
medium
to large colony: one Capnella sp.
Invertebrates:
five Trochus sp. snails
Water flow:
slow flow from an Ampmaster 2600™ centrifugal pump
(Dolphin Aquarium and Pet Products, Inc.)
Lighting:
four 48" 110 watt VHO fluorescent bulbs (two actinic,
two daylight - URI, Inc.)
Tank 5
Tank 6
Size:
30-gallon
Substrates:
7.5 cm medium grain aragonite, small amount of live
rock rubble (>25 kg)
Fish:
none
Corals:
none
Invertebrates:
five Trochus sp. snails
Water flow:
slow flow from an Ampmaster 2600™ centrifugal pump
(Dolphin Aquarium and Pet Products, Inc.)
Lighting:
four 48" 110 watt VHO fluorescent bulbs (two actinic,
two daylight - URI, Inc.)
Tank 6
Tank 7
Size:
30-gallon
Substrates:
7.5 cm medium grain aragonite, live rock (>10
kg)
Fish:
Chaetodon rostratus
Corals:
none
Invertebrates:
five Trochus sp. snails, 50 Nassarius sp.
snails
Water flow:
slow flow from an Ampmaster 2600™ centrifugal pump
(Dolphin Aquarium and Pet Products, Inc.)
Lighting:
65 watt power compact fluorescent 6500K (Lights of America,
Inc.)
Tank 7
|
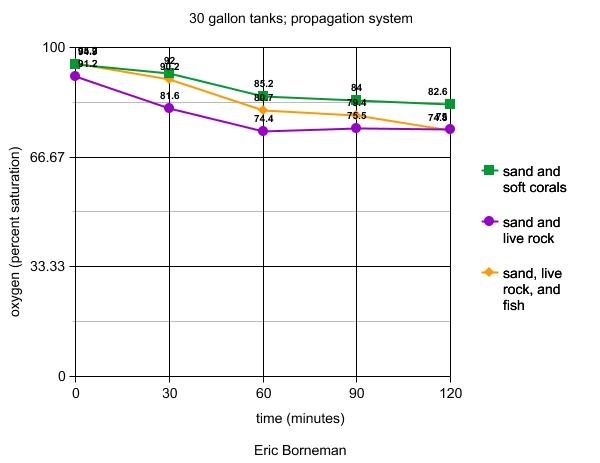
Table 7. Respiration of various communities and
their effect on oxygen concentration in three 30-gallon
seawater aquariums. All tanks were unstirred and unlit
during the experimental period.
|
Discussion
The tanks tested in this article show
some interesting and unexpected results. In the control tanks
with sterile water, a small initial peak occurred in both
samples, despite being sealed from outside air. This probably
resulted from the equilibration of existing chamber air (21%
oxygen) with the seawater since only half the desiccation
chamber was actually filled with sample water. The later slight
decline may be due to slight warming of the water sample from
the motor of the stir plate. Differences in the curves are,
as expected, statistically insignificant.
In the first experimental tank, the variations are slight
and the curves are statistically insignificant (Borneman,
in preparation). Any variations between the lines are likely
due to the reasons mentioned above, to the order in which
each condition occurred and to stochastic variations that
are nearly impossible to adequately control in multiple tests
involving numerous variables, non-sterile conditions, and
environmental exposure in a working laboratory.
The second and third experiments produced surprising results,
especially in terms of the variables' effects in non-hypoxic
tanks (next article). This set of experiments was recently
performed after Martin Moe, Julian Sprung and other attendants
at IMAC suggested that water movement and airstones, in particular,
might play a role in oxygenating water that was already reduced
in that gas. My data at the time had explored the role of
these variables only on various tanks or chambers with significantly
higher "normoxic" water conditions. In the case
of hypoxic water, such as might be found at night or in the
event of power failures, the results clearly show that airstones,
water pumps and skimmers are quite effective in rapidly raising
the oxygen level of both plain seawater and small reef aquariums.
Their effect on larger tanks, however, remains minimal and,
again, will be shown next month.
One finding which conflicts with previously explored data
presented at IMAC is the ability of photosynthesis to rapidly
raise oxygen levels in hypoxic seawater only. I will present
conflicting data and potential reasons for the discrepancy
in next month's article. It is also notable that a small piece
of coralline algae-covered live rock with a smallish colony
of Protopalythoa produced as much oxygen as a medium
sized tangle of Chaetomorpha sp. algae. In contrast,
stagnant diffusion across the surface of the test tank's water
was very ineffective in raising oxygen levels, and although
the test was discontinued after one hour, I believe it unlikely
that oxygen levels would approach the baseline levels for
a very long time. Part of this result might be due to the
test aquarium's surface area-to-volume ratio, which was much
lower than that of other tanks I had tested and found to equilibrate
much faster, even without circulation. Another factor that
I will examine in more detail is the PAR level to which the
algae were exposed (much lower in this experiment than the
previous data presented at IMAC). These comparisons and statistics
will also be provided in the next article.
In terms of the ten-gallon laboratory reef tanks, and despite
their lack of fish, it is apparent that supersaturation of
oxygen never occurs, although it is close to 100% in one case.
I attribute this, as a contrast to other tanks that will be
described next month, to the low irradiance provided by a
single 18" fluorescent fixture not allowing for maximum
photosynthesis, although a relatively high saturation in excess
of 90% is testimony to the relatively low light levels capable
of maintaining many corals and, together with a single water
flow device, far outpacing the respiratory demands of a reasonable
biomass of bacteria, algae and invertebrate species.
The fourth experiment examines the effects of light and darkness
without the variable of water motion. In both tanks that are
similar in many ways with some species differences and a lack
of a skimmer on one tank, the effects are expected and the
same in nature. Light alone in normoxic water provides enough
oxygen for the community to maintain stable oxygen levels,
while the community's respiration in darkness causes oxygen
levels to drop. Tank 2 displayed a slight increase in oxygen
in the lack of water motion, which corresponds with a similar
finding under high illumination of Chaetomorpha in
unstirred conditions that will be discussed next month.
The fifth experiment is similar in nature to the experiment
above except that its communities are distinct, the tank volumes
are tripled and the surface area-to-volume ratio is much greater.
Furthermore, a less precipitous drop in terms of the shape
of the curves, or even a leveling off in the case of the sand/live
rock and the sand/soft coral tanks, is interesting. Because
this system is exposed to ambient room light in my sunroom,
despite this period being considered "night" as
the system is set on a "reverse daylight" cycle,
there may have been enough light present for some photosynthetically
efficient organisms to produce some oxygen, leading only to
a slightly lower P:R compensation point in these tanks than
occurs when artificially illuminated. Alternatively, a continuing
decline might have occurred had the test period been carried
out over a longer duration.
Next month will conclude this series of articles, and I will
show the results of other experimental tanks, shipping containers,
and the dynamics of oxygen over normal and unmanipulated conditions.
Finally, I will prepare graphs showing and explaining the
various contributions of the variables tested in terms of
their effects on the communities found in reef aquaria. I
will produce several new sets of data, some of which were
presented at IMAC, but will concentrate on how the major variables
affecting oxygen in tanks play into an "average"
aquarium running normally. I will also tease apart each variable
and compare its relative importance under normoxic and hypoxic
conditions. Finally, I will conclude next month's article
with suggestions as to how to maintain higher oxygen levels
in tanks with very low P:R ratios (i.e. heavily stocked tanks,
tanks with high fish loads, tanks with low water flow or those
with low surface area-to-volume ratios) and consider the implications
of shipping livestock that must endure long travel times in
small water volumes.
|


)
)
)
)
)