|
"A thing of beauty is a joy forever:
its loveliness increases; it will never pass into nothingness."
John Keats
|
Figure 1. A colorful and truly beautiful temperate
aeolid nudibranch, Phidiana crassicornis. As
with all mortal things, unlike the beauty of Keats,
it will eventually die and pass into nothingness.
|
To Send a Message
One of the eternal
questions that has transfixed humans, probably since they
could think of such things, is, "What is beauty?"
We all "know" what beauty is, but it is really impossible
to come up with a definition that satisfies everyone. Perhaps
a more interesting question is, "Why is beauty?"
There are all sorts of reasons that we might perceive something
as being beautiful. Setting aside the biological reasons for
the perception of beauty within our species, people generally
define beauty as a combination of attributes that elicits
a pleasurable response. From the aspect of an evolutionary
biologist, many of the reasons given for the perception of
beauty devolve into the mechanical and, sometimes, arcane
world of perception of signals given by one species to another.
Individuals within one species often wish to send signals
to individuals of other species or just to the world in general.
Perhaps the easiest of these signals to understand are those
sent by one species to enlist the aid of another. We are all
familiar with the beautiful color of many flowers, but many
people don't realize that these colors are the plant's way
of sending a message to some animal to come and help the plant
reproduce.
Flowers' colors are often specific messages to particular
groups of animals to tell them that the animals can get a
reward, generally food in the form of nectar (sugar water)
or pollen (rich in proteins), by visiting that flower. When
the pollinator visits successive flowers, it can transfer
pollen from one plant to another, thereby helping the plant
reproduce. Neither the plant nor the pollinator "thinks"
of this process in this way, of course. Plants have no nervous
system and can't think, and all the pollinator is after is
food. However, the transfer of pollen is not incidental. As
the plant with the best signals gets the most visits from
pollinators, and this can lead to more offspring, the whole
system forms a positive feedback loop under the control of
natural selection. Once the system gets going, it can get
fine-tuned for specific pollinators; and if the signal for
a specific pollinator is sufficiently common, some plants
may develop a way to cheat. They give the signal to the pollinator,
but don't offer the reward. Pollinators may still visit these
plants and transfer pollen, but the plant doesn't have to
expend the energy to produce the reward.
|
Figure 2. Similar signals, different rewards.
Left: The fairy slipper orchid, Calypso
bulbosa, rewards pollinators, such as bumblebees,
with pollen. Right: The color pattern and
shape of the lady slipper orchid, Cypripedium
parviflorum, seem to promise a reward; instead,
the flower traps pollinators in its pouchlike
"lip." When they crawl out they get
dusted with pollen. If they are then tricked into
another lady slipper, the pollen is transferred.
|
|
Not all signals are rewards, either; in our terrestrial environment
we are all aware of the way certain animals signal to other
species to tell them to avoid attacking or even getting close.
Some animal signals are active, such as the facial expressions
of a dog that feels threatened and cornered; but many are
also passive, such as the coloration of patterns of Pepe
Le Pew and his handsome, but malodorous, kin,
and those of the yellow
jacket wasps. I would bet that very few people who read
this have ever been tempted to reach out and grab a large
wasp. The avoidance of such patterns is instinctual in many
animals and probably is in humans as well; but in addition,
if someone does reach out to hassle said flying insect, he
is liable to discover just how intensely certain insects can
activate parts of the mammalian nervous system devoted to
signaling pain. The learning experience that accrues from
such an initial miscommunication ensures that henceforth the
insect's message of, "I am dangerous, leave me alone!"
is well and clearly understood.
Unlike many other terrestrial ethological situations, this
type of signaling has exact analogues in the marine realm.
In fact, as animals in the seas are much more diverse in body
form, and have been evolving for much longer, than those on
the land, many more examples of these behaviors exist in the
oceans. The use of color or color patterns to signal or warn
of an interaction's potentially unpleasant outcome is found
throughout marine environments where there is enough light
to see the colors. Coloring to signal potential danger is
referred to as aposematic coloration and is almost ubiquitous.
Figure 3. Warning coloration in dorid nudibranchs.
Left: An individual of Phyllidia varicosa, a
coral reef nudibranch that is so toxic that the death of a
single individual in an aquarium has caused fish mortalities.
Right: Triopha catalinae, a toxic temperate
nudibranch that lives in turbid waters. The brilliant white
and contrasting orange coloration ensures a visible warning
signal even in murky seas.
Having a color pattern of any sort implies the production,
utilization and storage of pigments. Pigments are often complicated
chemicals that may be metabolically "expensive"
to produce and maintain. Thus, there is a cost of having "color"
of any sort, but having a protective coloration is better
than being dinner. Consequently, animals without a way of
deterring their potential predators are often camouflaged.
Another obvious, but oft forgotten, implication of either
a warning or camouflage coloration are that the predator can
see. As a corollary of these factors, animals that are preyed
upon by predators that don't have eyes, such as those eaten
by sea stars, are not aposematically colored, but might possess
tactile camouflage, such as the covering of shells and debris
that many sea urchins "wear."
Most predators are not indiscriminate in their choice of
prey, and as a result, warning colors are often tailored to
specific predators; we humans tend to forget that most animals
are not as visually well-endowed as we are. While many animals
can detect changes in light intensity, relatively few of these
animals can detect images. Most animals with eyes have simple
eyecup photoreceptors that can detect only changes in light
intensity or direction. Visually capable predators in the
marine environment are limited to four animal groups: the
annelid worms, the cephalopod mollusks, the crustaceans and
the vertebrates.
If I define "good vision" as the ability to form
and process images, then good vision is not widespread in
the annelids; only a few of the polychaetes have well-developed
eyes, but those that do have them, such as some pelagic swimming
predators, have excellent vision. Good vision is obviously
found in the octopods and squids, but their distant cousins,
the chambered nautiluses, have only "pinhole" camera
eyes that are not very good visual organs. Many crustaceans
have compound eyes, which may be very good image forming eyes,
albeit with a different type of image than we would see. Within
the marine vertebrates virtually all animals have relatively
good vision, and virtually all of them are predatory, if not
on animals then on various algae and protists. Some people
may think that predators eat only animals, but this is, of
course, poppycock. Predator-prey interactions as a subset
of ecology or ethology encompass all potential predators along
with all potential prey.
As I have described, visual predators and their prey are
often involved in a co-evolutionary dance. Natural selection
fine-tunes predators by promoting attributes such as more
visual acuity and interpretative capability. An interesting
corollary to this is, all other things being equal, predators
are usually more intelligent than their prey. As a professor
of mine once put it, rephrased so that the censors won't bleep
it out, all a prey animal has to do to be successful is "breed
like a bunny" and run like hell. That is an over simplification,
of course, to make a point. But, dogs and cats make far more
interactive pets than do rabbits or sheep. In aquaria, predatory
animals such as fishes have a whale of a lot more personality
than do their prey such as sponges, algae, or corals; this
even may be said for fishes that eat other fishes, for example,
lion fishes are often more interesting animals than are the
fish that they may eat. The co-evolutionary dance works on
the prey as well, though. They can develop means of hiding
in plain sight, such as camouflage, or by behavioral attributes
such as nocturnal foraging. Or… they may become dangerous
to eat.
Nudibranchs = Mobile Poison Packages
All aquarists are familiar, in some
regards, with snails. Snails are mollusks with a very peculiar
internal anatomy; their guts are twisted 180º in relation
to their head/foot region relative to all other mollusks.
This means that in most snails, their anus opens not at the
animals' rear, but rather just behind their head on what would
be their right shoulder, if they had shoulders. This internal
anatomical twist, and nothing else, defines
what a snail is. Shelled snails are rather well-protected
from predation. The molluscan shell probably evolved as a
protective covering of some ancient worm, and it is supremely
well-adapted for its function. One of the more interesting
aspects of snail biology is that most snails can live quite
well if their shell is removed. It happens occasionally even
in nature. During the course of my doctoral research I collected
and examined a skosh over 15,000 individual snails from subtidal
and intertidal environments. Two of them were effectively
naked. They had just a small remnant of their shell remaining,
having lost the majority of the shell to breakage or erosion.
Other researchers have noticed the same thing. In nature these
snails eventually perish from predation or simple mechanical
damage to their unprotected tissues, but until that happens
they can live an effectively normal life.
If such naked animals had a way to protect themselves, they
presumably could live and persist. Many snails and other marine
animals accumulate materials from their foods in their tissues.
This is particularly true of toxic materials. Indigestible
toxic materials often are "physically sequestered,"
that is, enclosed in some sort of protective container, often
membrane bound vesicles, which effectively seals them off
from the body's metabolism. These materials, such as the heavy
metals laughingly known in the aquarium hobby as "beneficial
trace elements," are generally so poisonous that the
animal can't excrete them without destroying its kidneys Both
organic and inorganic poisons may be found in many marine
animals, including snails, either enclosed in vesicles, or
maintained in an insoluble and therefore non-toxic form, or
incorporated in tissues in vacuoles wherein the internal conditions
alter the toxic chemical to make it non-toxic. These various
forms of detoxification are common in today's mollusks, and
given the antiquity of the group, were probably also found
in at least some of the ancient forms. Knowing that, it is
not too hard to envision a scenario in which some particular
ancient snail lost its shell, through some sort of genetic
mutation. All of the animals in the taxonomic order Nudibranchia
develop a
shell during their larval growth. And all of them drop
the shell from their bodies when they metamorphose from the
larval phase to the juvenile form. Given that all nudibranchs
do this, and given that they all do it in the same manner
at the same time in their lives, it is likely that such a
property was inherited from a common ancestor.
This "shell shedding" could be the result of a
single mutation that happened in the ancient seas. It appears
that the nudibranchs separated from their shells and in doing
so, from their ancestral group of snails sometime in the middle
Paleozoic era, probably around 350 million years ago. The
common ancestor to all of today's nudibranchs was likely a
shelled animal not terribly dissimilar to some of the grazing
snails found in today's reef aquaria. This animal was carnivorous,
and probably grazed on some sort of sponge or other toxic
animal and in some manner it probably sequestered the poisons
from its food in its tissues.
Once the nudibranchs separated from the main line of shelled
snail evolution, they also diversified. Such small animals
must have appeared as a snack to the predators of the time,
and there were some fearsome benthic marine predators during
that period. If the ancestral nudibranchs were toxic, it would
be to their distinct advantage to be aposematically colored.
This would drastically reduce their mortality as predators
could learn to avoid them. The more distinctively colored,
the more rapidly would the predators learn to snack elsewhere.
It appears that being toxic and advertising that fact became
a generalized nudibranch characteristic; today very few nudibranchs
appear to be palatable to any other marine predators. Interestingly,
those that are palatable are almost always camouflaged.
Spicy Slugs
During the winter,
the native inhabitants of St. Lawrence Island, one of the
Pribilof Islands in the Bering Sea, sometimes collect, chop
up and eat pieces of the large plumose anemone, Metridium
giganteum, which can be found in the lower intertidal
zones of that area. Whilst teaching in Alaska eons ago, one
of my students from that area told me that it tastes "spicy,
kind of like hot peppers." This "spiciness"
was, of course, due to the stinging capsules or nematocysts
found in the anemones' epidermis. These nematocysts were discharging
into the tongue's epithelia, giving the feeling of spiciness
(and adding a note, with thanks, from Mr. Borneman when he
reviewed this article, "Capsicum, or the chemical found
in hot peppers, incidentally, fits almost perfectly into pain
receptors.") Interesting the things we humans eat for
"spice." Metridium is an anemone that feeds
on planktonic particulate materials and doesn't have particularly
venomous nematocysts, fortunately. Other common anemones in
the area do have much more potent nematocysts, but the natives
of the region didn't eat those, probably because they learned
long ago that their "spiciness" could be extremely
unpleasant or even lethal. Such experiences are lost in antiquity,
but the spiciness of the anemone gives an indication of the
effects of nematocysts on a predator, albeit a most unlikely
one. If the predator were far smaller, the nematocysts' effects
would be more extreme and dangerous.
Somewhere very far back in time, however, some small predatory
sea slug developed a way to eat some of the cnidarians of
its time. The snail gut is not like that of a human. Snails
typically eat food by rasping it into very small pieces and
ingesting them. Once the food has been eaten and is in the
stomach, the fine food particles are sorted by weight, density
and size. My guess is that some snails that ate cnidarians
managed to have the right combination of gut chemistry and
prey items so that most of the nematocysts were not discharged
during feeding. Snails also do not digest food the way that
vertebrates do, in the gut cavity. Instead, the fine - microscopically
fine - particles that constitute the snail's ingested food
are only sorted in the gut and those that fall into the right
range of size and density are moved into some large glandular
structures on either side of the gut. These are called "digestive
glands" or sometimes - and very inappropriately - livers.
These glands are a collection of very small tubules surrounded
by masses of digestive cells. The cells individually ingest
food particles and digest them internally. Excess nutrients
are liberated to the blood, bathing these cells, and are then
transported to the rest of the animal. Indigestible particles
are moved out of the digestive gland back to the stomach where
they are shunted to ciliary tracts that take them to the intestine
and then out of the animal.
Digestion of a nematocyst or something similar would be dangerous,
and the ancestral nudibranch probably simply held these for
a brief period, a few minutes to a couple of hours, in its
digestive gland and then moved them back to the stomach to
be excreted as feces. However, over time and in some particular
species, the nematocysts were probably maintained for some
longer period in the digestive gland. If such an animal were
to be eaten by a predator, these nematocysts would be prepositioned
to discharge if the nudibranch's tissues were damaged. Depending
upon the prey initially eaten by the slug, such a nematocyst
discharge could give the predator a really unpleasant sensation.
If that particular species of slug was distinctively marked,
the predator could easily learn to avoid such a nasty-tasting
food item. This could have been the beginning of the coevolution
of the predator and its prey, as the distinctive coloration
could easily change to become aposematic coloration.
Today, many people find nudibranchs of all types to be among
the most beautiful of all animals. They are often vividly
and brilliantly colored with striking color patterns that
we find aesthetically pleasing; color patterns that many marine
animals note as a warning to avoid. While the most colorful
nudibranchs are those in the suborder Doridacea, which eat
toxic sponges and bryozoans and use striking colors and color
patterns as warning signals, many nudibranchs, particularly
the larger ones in the suborder Aeolidacea are also considered
to be very beautiful. These animals, the aeolids, are specialized
to eat cnidarians. They store the eaten, but undigested, nematocysts
of their prey in pouches extending up from their backs, presumably
as predator deterrence.
Aeolids, the Prey that Bites Back
Figure 4. A small Flabellina showing the type
of warning coloration typical
of aeolid nudibranchs.
Most snails are fairly small animals
and many aeolid nudibranchs epitomize this. It is a really
large aeolid that is over a couple of centimeters long, and
many of them reach adulthood while still less than a centimeter
long. Aquarists and the general public often seem to conceptually
link small size with simplicity of design. That is a mistake
with these slugs, however, as they have a complexity of internal
structure that is truly impressive. There are probably less
than a thousand scientifically described species of aeolids,
although it is highly likely that a great many more remain
to be described.
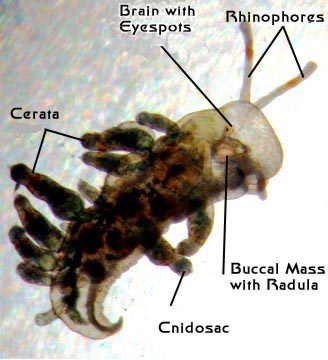
Aeolids are nudibranchs, which means that they are snails
that have lost the shell they had as larvae. Once they pass
out of the larval stage they have no shell or remnant of one.
Being snails, they have, for the most part, a rather straightforward
internal morphology. They have a typical snail's nervous system
consisting of several ganglia surrounding the throat. This
ring of ganglia constitutes a reasonably large brain, relative
to the size of the animal. Nerve tracts or cords run posteriorly
through the body, and there may also be subsidiary nerve cell
aggregations in the posterior part of the body. Their sensory
structures are a bit odd. Unlike most snails, they lack eyes
on the surface of the body, but their eyespots are well developed.
These sit right on the dorsal-most ganglia that surround the
foregut under several layers of tissues. All nudibranchs have
a pair of sensory tentacles that arise from the region of
the body that would be the top of the head, if they had a
distinct head, which they don't. These tentacles are called
"rhinophores," and they contain sensory structures
sensitive to current flows, dissolved chemicals, and touch.
Sort of "all purpose sensors," these structures
have been referred to as "world sensors."
Figure 5. An adult individual of a species of Eubranchus.
The animal was about 3 mm (1/8th
inch) long. This small
nudibranch eats hydroids.
They have a well-developed heart consisting
of one auricle and one ventricle. Blood, containing hemocyanin,
a copper-based respiratory pigment, is pumped from the heart
through vessels toward a couple of bodily regions. These vessels
soon end, and the blood flows throughout the body, bathing
the internal organs. This type of circulatory system, lacking
most arteries, capillaries or veins is called an "open"
system. Although it lacks vessels, the blood flow is neither
indiscriminate nor haphazard; the blood flows through gaps
in the tissues that direct it in a precise pattern. Eventually
the blood passes through a kidney and then to the heart to
complete the circuit. Gas exchange in aeolids takes place
over their body's whole surface, but especially in the frill-like
extensions, called cerata (singular = ceras), which extend
up from the animal's back.
They are simultaneous hermaphrodites and their reproductive
system's structure varies significantly among the various
subgroups of aeolids. Some species' reproductive plumbing
is among the most complex, if not the most complex, in the
animal kingdom, containing a male system, a female system,
and a tertiary system designed to store and hold sperm, often
for several days to a week between copulation and spawning.
Even though they are hermaphrodites, they are not self-fertile,
so two animals are needed to create viable offspring. Unlike
the dorid nudibranchs, which will copulate for hours on end,
many aeolids have very short copulatory periods; Phidiana
(=Hermissenda) crassicornis, a common temperate
species used in neurobiological research, can complete simultaneous
and reciprocal (remember, they are hermaphrodites) copulation
in under one second. After copulation, sometimes weeks after,
aeolids typically deposit a gelatinous egg mass containing
"eggs," which actually are developing embryos. These
typically hatch from the egg mass and spend from a week to
a few months in the plankton, feeding and growing. During
this period, they have a typical larval snail shell. When
they have developed sufficiently, they settle from the plankton,
shed the shell and start crawling around looking for food.
If this were the only developmental pattern, these animals
would not be any real problem for aquarists, as the larvae
would seldom survive their larval period due to the scarcity
of plankton upon which to feed. However, a few species - and
perhaps more than a few species - have a rapid, totally benthic
development that occurs in the egg capsule mass or, alternatively,
have a larval period of a day or less. These species can complete
their life cycle in aquaria, and some of them can become quite
serious nuisances.
In addition to their complex reproductive systems, they have
a gut system that has some of the strangest modifications
seen in any gut in any animal. The front end of the gut is
normal; for all intents and purposes it is indistinguishable
from any other small snail's gut. As is the norm for snails,
they have a well-developed radula. In this case, it is not
a rasping organ and the number of radular teeth has been reduced
to only a few, sometimes only a single tooth
in each row. Aeolids typically also have jaws that help in
slicing off prey. Aeolids are generally specialized to eat
cnidarians such as corals, sea anemones and hydroids, although
a few oddballs eat other things.
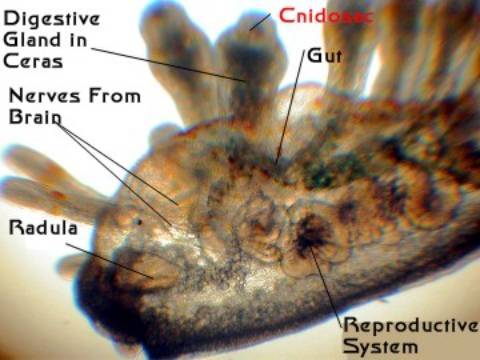
Figure 6. The same Eubranchus individual as
in Figure 5, showing the relationship
of some gut structures, cerata and cnidosac.
During feeding the prey tissues are cut into small pieces
and swallowed. They then pass into the stomach through a short
esophagus. So far, so good; everything is "up to the
molluscan standard." Once in the stomach, the food is
sorted and digestible foods are sent to parts of the digestive
gland where cells lining the tubules ingest and digest them.
With nematocysts, however, a different set of events takes
place. They are sent into finger-shaped, blind-end tubules
which extend up into the cerata. Each ceras contains one digestive
gland duct. At the end of each of these tubules is a specialized
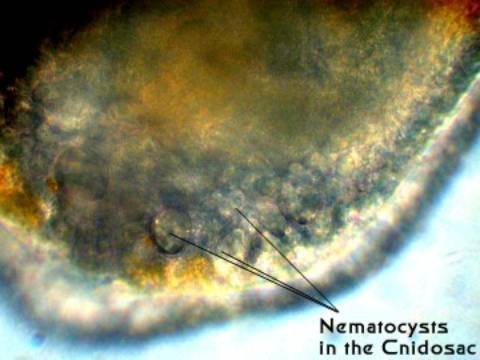
sac or pouch, called a "cnidosac."
This is often highly pigmented; generally, it is colored with
brilliant white pigment. Presumably, secretions from the inner
walls of the cnidosac maintain the nematocysts in a fully
functional condition. Other nudibranchs, such as the dendronotids,
also eat cnidarians, but only the aeolids keep the nematocysts
within their bodies as functional structures. Aeolids are
not toxic in the classic sense. They don't have chemicals
in their bodies that deter predation. However, their defenses
are also effective. They take internal structures from their
prey and use those as defenses against their predators. If
something like this existed on land, it would be as if a coyote
ate a porcupine, quills and all, and then its digestive system
deposited the quills in intestinal pouches growing out of
its back to protect it against wolves. Needless to say, this
whimsical terrestrial example doesn't occur. But, the marine
version does, and it is found in all seas, as aeolid nudibranchs
are common in most shallow water marine environments. Although
this seems to be an arcane and very unlikely way to protect
against predation, the sting of these nematocysts may be very
serious. Human
injuries from contact with aeolid cerata are not uncommon.
Instances
of very serious injuries resulting from contact with the nematocysts
concentrated in aeolids have been recorded from aquaria.
Figure 7. Nematocysts in the cnidosac of the Eubranchus
individual pictured previously.
Aeolid coloration is somewhat related to size. Large species
are often brightly colored with aposematic coloration. Smaller
ones tend to have color patterns that result in aposematic
contrast patterns dominated by dark lines on a white background.
Even the most colorful patterns are difficult for us to interpret,
though. Not all marine visual predators see colors, and those
that do may not see colors the same as we do. Nevertheless,
the larger aeolids seem to use a larger palette of colors
for their patterns, while the smaller ones are more "black
and white."
Aeolids in Aquaria
With only a couple
of exceptions, aeolids should not be welcomed with open arms
by aquarists. They all eat cnidarians, and many of them are
specialized to eat some species of the decorative livestock
that aquarists keep. Numerous species have found their way
into aquaria from time to time. Generally, the solution to
maintaining these animals is simply to remove them. Solitary
hitchhikers are seldom able to set up or maintain a breeding
population. However, there are some exceptions. Some, typically
small, species can pass through their entire life cycle in
a reef tank. These particular nudibranchs pass through most
or the entire larval period within the egg mass, and do not
need to feed in the plankton. Consequently, they can proliferate
in an aquarium. As these nudibranchs are specialized in diet,
they have picked up the common names of "Montipora-eating
nudibranch" or "zoanthid-eating nudibranch."
It is likely that there are several species in each group,
and controlling them is difficult. Being aeolids, few animals
will eat them, and being small and mobile they can spread
throughout a tank and infest it before being noticed. Among
their few predators are some other slugs (see here
for some movies of them being eaten; note how well the cerata
repel the potential predator, Pleurobranchaea, and
are ineffective against the other, Navanax). Interestingly
enough, these other slugs lack eyes, so any aposematic coloration
is "wasted" on them. Dips of various sorts may be
used to control them in aquaria, but these vary in efficacy.
As the converse of these problem aeolids are several
other small species, all sold under the "umbrella"
name of "Berghia."
These nudibranchs are bred and sold specifically to prey upon
the pest anemone species in the genus Aiptasia. Large
aeolids are often strikingly beautiful animals, but they are
not reef aquarium safe. Smaller aeolids tend not to be as
highly colored, and some of them are useful in aquaria, but
the majority of these are also not animals most hobbyists
would want to try to maintain in their aquaria. As with numerous
other highly colored marine animals, one might wish to recall
the aphorism that "Beauty is in the eye of the beholder."
|

)


)
)