|
Life With A Twist
Success.
What is success, and how do you measure it? We all know that
there are different definitions of success in the various
fields of human endeavor. For some people, success is more-or-less
equated to power. For others, it may mean being content with
their lot in life. For others, it may translate in to how
much good they have done for their fellow humans. It is obvious
that the definition of success is arbitrary and depends on
the individual and the context of the discussion.
To some extent, the same holds true when discussing the biological
world. Within a given group or taxon, success may equate to
the number of species found throughout the world. Within other
contexts, it may simply mean survival. If life evolved but
once, then we are all co-descendants of that one event, and
any living organism is monument to some sort of successful
longevity. By most estimates, easily 99.999% of all the different
types of organisms that have ever lived have come and gone
and are extinct (Tasch, 1973; Raup, 1976a,b). Success, in
this case, would simply mean persistence.
Other definitions of success, though, relate
to the number of evolutionary descendants. By this measure,
one of the greatest success stories within that portion of
life we call the "Animal Kingdom" is the group termed
by taxonomists, the Class Gastropoda of the Phylum Mollusca.
Commonly known as "snails," only the great arthropod
group we describe as insects has a greater number of scientifically
described species. Estimates of the number of living snail
species range from well over 100,000 to somewhat less than
30,000. The wide latitude in these numbers indicates the large
degree of uncertainty due to a simple lack of knowledge. No
matter how you cut it, however, there are a LOT of snail species.
Considering this vast number of species, one of the more interesting
questions a person could ask is, "Why? What is it about
being a snail that confers such a good chance of evolutionary
success?" The answer may be in the way snails have partitioned
the world into functional units, and to a very great extent
this is based on their own structural peculiarities.
Looking beyond the basic differences between
mollusks and arthropods, some obvious factors differentiate
the degrees of variance between the snails and the insects.
One of the interesting oddities of animal diversity is that
within the huge group of insects, they really do not look
all that different. After having taken a really good look
at an insect, it is unlikely that an adult insect would ever
be confused with an adult animal of any other group. Not so
with the snails. Only one characteristic separates snails
from all other groups of mollusks, and that is that snails
undergo torsion. Torsion is an internal twist that generally
occurs late in the snail's larval life. During torsion, the
animal's viscera behind the head and above the foot rotate
180 degrees, bringing the anus to a position pretty much right
above the head. Now, this process is bizarre in the extreme,
and is probably related to the animal’s locomotion and
the way it must carry its shell. Incidentally, torsion does
not equal coiling. All snails have undergone torsion whether
or not they have coiled or uncoiled shells, or whether or
not they even have shells at all. However, the point is that
the only thing that absolutely defines snails is internal
and invisible to the casual observer. Furthermore, unlike
the insects, whose morphology is confined and defined by their
exoskeleton, the snails have notoriously variable shapes.
They can look like just about anything.
Snails come in a wide variety of shapes
and forms; however, there appears to be only a finite number
of basic ways to make a snail shell (Raup, 1966; Raup &
Graus, 1972). Within such groupings of snails with similar
shells, the animals often exhibit some common attributes.
Although grades of structure often include groups of closely
related animals, kinship is not implied by their structural
similarity. Many groups look alike and have similar structures
due to convergent evolution. In effect they get to about the
same endpoint, but by different pathways, and from different
origins.
One such structural level is called the
"archaeogastropod" grade of structure. These are
the snails that have shells similar in some regards to the
earliest snails in the fossil record. Anywhere from a few
hundred to a few thousand species can be termed archaeogastropods.
They are important to aquarists as this group includes most
of the grazing snails that are used to control algae. These
are animals such as the abalones,
keyhole limpets, many other limpets, and the "turbo"
grazers such as animals in the genera Astraea,
Turbo, Trochus, and Tectus. With
all of that, however, the archaeogastropod structure is relatively
consistent and most people can recognize one once they have
seen a few of them (Abbott, 1974; Abbott and Haderlie, 1980).
There are two other large groups of marine
shelled snails, each of which also comprises animals with
distinct basic appearance. One of these, the "neogastropods,"
comprises the largest array of snail species. These are the
whelks, the venomous snails, such as Conus, and their
kin and literally tens of thousands of species are put into
this grouping. As in the insects, however, this group’s
diversity is not manifested in widely differing shapes and
structures. Rather, they have diverse internal specializations
reflecting their modes of predation. Most of these snails
are active predators, and their immense diversity is probably
related more to their ability to specialize on specific prey
than to their innovative external forms.
The remaining structural group has been
referred to as the "meso- (or middle)" gastropods,
because in many ways they are morphologically intermediate
between the archaeogastropods and the neogastropods. Evolution
and development, in general, and gastropod evolution and development,
in particular, may be viewed as a sequence of advances, each
overcoming an obstacle that allowed for the exploitation of
new habitats or food sources. Although the archaeogastropods
have similar shells, the many different species possess almost
as many different combinations of gills and excretory systems.
Consequently, hidden within the similarity of shell shapes
are a great many different arrangements of internal plumbing.
In snails, unlike vertebrates, the respiratory and excretory
systems are integrated to a great degree; blood flows directly
from the gill into the kidney and then to the heart. This
means that the heart is pumping not only freshly oxygenated,
but also freshly cleansed, blood to the tissues. However,
there are many ways for the respective plumbing systems to
be interconnected and related. Most of these appear to have
some deficiencies and are not very efficient.
At about the same time that reptiles started
to become common on land, gastropods that were recognizable
as mesogastropods started to occur in the fossil record. This
marked the beginning of one of the greatest documented evolutionary
radiations known. Archaeogastropods, in general, are limited
to crawling and feeding on hard substrates. The mesogastropod
grade of respiratory and excretory plumbing was apparently
all that was needed to start to exploit most of the other
marine habitats and methods of feeding.
So, while all of the archaeogastropods have a fairly limited
and simple repertoire of shapes, all related to grazing on
hard substrata, the mesogastropods literally exploded into
hundreds of shapes, sizes, and habitats. Today, the mesogastropod
grade of structure is found in about 200 distinct and not
closely related lineages. These include such diverse groups
as cowries, predatory moon snails, mud-dwelling cerithiids,
eulimids which are parasitic inside of sea cucumbers, and,
of course, today's main course of this vast gastropod meal,
the worm shells or vermetid gastropods (Abbott, 1974).
How They Do It
Worm
shells are so-called because their shells look superficially
like the tubes of the calcareous feather-duster tube worms
known as serpulids. The Serpulids’ head is modified
to act as a filter-feeding organ. As with all feather duster
worms, this filter-feeding organ is constructed of a lot of
finely and pinnately branched tentacles, giving the appearance
of tiny feathers on a feather duster. This characteristic
structure, however, has nothing to do with their shells, which
are often simple straight or coiled tubes. Because the animals
construct a small tube when they are small that gets larger
as they grow, the tube has the shape of a long, narrow cone
that is straight or meanders across the substrate. However,
some of them are coiled around their base, very much in the
manner of a coiled snail. On the other hand, worm snails start
out life as a small, rather normal-looking, snail with a coiled
shell, albeit of only one or two whorls. After a short period
of free-living life, they cement their shell to a hard substrate.
As they grow, the shells may coil or meander over the substrate
producing a tube that looks quite similar to a serpulid tube
worm shell. However, the tube worm produces a shell that is
generally dull-surfaced on both the inside and outside, while
the snail's tube is glossy inside. The worm's tube begins
as a simple tubular chamber containing the recently settled
juvenile worm. The worm tubes are generally white, although
they often become colored with coralline algae or other epibiotic
growth. In contrast, the small snails start out with larval
shells that are tightly and spirally coiled. After the juvenile
snail cements itself to the substrate, its shell begins to
grow, generally in loose coils, at right angles to the spiral
of the larval shell (Keen, 1971). The shells can form quite
large entwined masses that are effectively impossible to separate,
containing dozens to thousands of snails. Enough snails may
become cemented together so that they may actually form reefs,
although these reefs are never particularly large.
 |
Figure 1. The shell of a small vermetid from my tank.
Note the several normal snail whorls
to the left. This whole animal was about ¼" from
top (left) to bottom (right).
Morphologically, of course, these animals have all the internal
characteristics that define snails. Unlike the worms, their
body is not divided into segments. They have undergone torsion,
which in their case is a decided advantage as it places the
anus at the front of the tube-shell. Consequently, they can
easily defecate undigested foods out of the shell. Most feather
duster worms have their anus at the back of the tube and have
special morphological adaptations, such as ciliated grooves
that serve to transfer their feces out of the tube.
The snails' tubes may be closed by a concave,
proteinaceous door or operculum. Reflecting their immobile
status, they have a reduced foot that is used mostly in feeding.
They possess a pair of relatively large tentacles on the foot,
each with an inner groove. A large mucus-producing gland is
located in the foot near the tentacles and discharges through
the tentacular grooves. Their gut is somewhat peculiar for
a snail in that the stomach contains a large rod of hardened
mucus called a "crystalline style." Crystalline
styles are more typical of bivalves, and contain digestive
enzymes (mostly enzymes that break down sugars) embedded in
the mucus. The style sits in a sac off the stomach and is
secreted at one end of that sac. Cilia in the sac and stomach
rotate the style at high speed (in some mollusks the style
can rotate at several hundred RPM). The rotating tip of the
rod is held against an abrasive area in the stomach, which
wears the tip off, liberating the enzymes and mixing them
with food that is brought into the stomach in a mucus strand.
This particular structure seems to be most commonly found
in herbivorous and plankton-feeding mollusks (Hyman 1967).
 |
Figure 2. Vermetus sp. photographed in about
1 m of water on the reef flat in Palau. The animal's aperture
was about 5 cm (2") in diameter. The brownish operculum
plugging the aperture and the mucous feeding strands are evident.
The feeding methodology of these animals is rather bizarre
and interesting. The animals use the mucous gland in their
foot to produce a large of amount of mucus. The mucus is extended
up into to the surrounding water by the tentacles on the feet
(Hyman, 1967; Kohn, 1983). The strands can extend quite some
distance depending on the water flow and the size of the animals.
In my aquarium, vermetids about 3 mm (1/8th inch) across can
project mucous strands over 60 mm (2.5 inches). I have seen
some large vermetids that were over 50 mm (2 inches) in diameter
on reef flats in Palau. The strands of mucus from these animals
extended over 2 m (about 6.5 feet).
Mucus is sticky, and planktonic materials adhere to it. After
a short time the animal "reels in" the strand with
its catch stuck to it and eats it. Some species have been
documented to feed together. When one individual starts to
put out mucus, all of its neighbors do too, producing a mucus
sheet that seems especially good at collecting plankton. Once
one individual starts to withdraw the strand, all of the contributors
do as well, and all get to share in the catch (Hyman, 1967).
This ciliary-mucous suspension-feeding isn't the vermetids’
only feeding mode, though. They also have been documented
to extend from the tube and catch small planktonic animals,
and they seem especially responsive to crustaceans (Hyman
1967). In aquaria, they are probably quite able to feed on
baby brine shrimp, as well as other small planktonic animals.
T'ank You For The Good Habitat
Vermetids
seem well-designed to reproduce in aquaria. Unlike most mobile
mesogastropods, they do not copulate. The males, however,
produce packets of sperm called "spermatophores"
which are transferred to the female's mucous nets by a pedal
tentacle, expelling the spermatophore into the water and "hoping"
she will catch it. This is not a forlorn hope; the animals’
gregarious nature often means that someone of the opposite
gender is nearby. The females collect the spermatophores and
store the sperm to fertilize their eggs. Embryos develop inside
the female’s tube and are maintained there until they
have passed through the larval stages and have metamorphosed
into little juvenile snails. They then leave the female and
crawl around briefly, usually for an hour or less, before
they cement themselves to a substrate (Strathmann, 1987).
Typically, the tubes’ apertures extend
upward, probably as an adaptation to facilitate spreading
of the feeding web. As the animals grow, they tend to erode
a hole through the side of the tube fastened to the substrate
and grow a new extension out of it; as they do so, they seal
off the old aperture with shell material. At the end of the
new extension, they construct another slightly larger vertical
extension with the aperture at its end (Keen, 1971).
Figure 3. Small vermetids about 3-4 mm (1/8th inch)
high photographed
in my system's refuge tank. They look like small calcareous
tube worms.
Vermetid
snails are relatively diverse; over a hundred species
have been described, and some of them are commonly found in
aquaria. Although several species are found occasionally in
our systems, generally entering on live rock, one variety
in particular may become very abundant, and be a serious nuisance
in some systems. This species, probably the most common, is
small, with a brown, reddish, or purple shell. Interestingly,
the animal is difficult to identify, although that has not
stopped numerous reef aquarists from doing so. It probably
is Spiroglyphus annulatus, which is a small vermetid
originally from the Caribbean. However, similar small species
live elsewhere in the world, and they all look pretty much
alike. It will probably take genetic testing to verify the
identity of our aquarium friends. Whatever species it is,
this particular one has small individuals. The tube seldom
is over one or two millimeters wide. The shells are typically
reddish or reddish-brown; sometimes they are even tinged with
violet. The animal forms a small, calcareous shell mound and
then sends up a short, three to five millimeter long, vertical
stalk. The upper edge of this tube may be razor sharp, and
may inflict rather nasty cuts. A few of these would be no
real problem; however, this animal reproduces very well in
marine aquaria. Left unchecked, it can reach populations of
over several thousand in a few months. They prefer high current
areas, and will infest and clog plumbing, significantly reducing
water flow. In severe infestations they can clog and shut
down pumps. The only solution in cases like these is physical
removal of the animals using whatever method is easiest (a
muriatic acid bath works well).
Fortunately, some fishes such as Copperband
butterfly fishes, seem to eat them, and some hermit crabs
will eat them as well. Eating these worm snails may well be
the only truly beneficial effect of hermit crabs in aquaria.
The larger vermetids found in reef tanks are probably in
the genera Dendropoma and Serpulorbis (Abbott
and Dance, 1982). They do not seem to reproduce well in our
systems and never obtain the plague proportions of their smaller
cousins. These larger species tend to enter our systems on
live rock or in coral, and are more interesting curiosities
than any kind of pest. For some reason the larger species
don't seem to proliferate as rapidly, though, and often remain
as relatively solitary animals. The larger species seem to
be more likely on Indo-Pacific live rock. A moderately large
vermetid in the genus Petaloconchus is common in the
Caribbean, and makes its way into aquaria now and then on
aquacultured live rock. Given the appropriate conditions it
is likely it will proliferate as well.
Figure 4. Several vermetids, possibly Dendropoma
sp., photographed in Palau, at a depth
of 15 m. These animals were about the diameter of a pencil.
The operculum and the
snail's tentacles are visible.
It is unlikely that even a large number
of vermetids is directly deleterious to any other aquarium
life. Sainsburys Offers the
freshest and popular products for weekly shopping! The mucus they produce may be used as food by many other
animals as well as by the producer. Large masses might produce
enough mucus to cause some local disruption in water currents
or they may foul some other animal, but the mucus is generally
very diffuse and most animals can easily remove it.
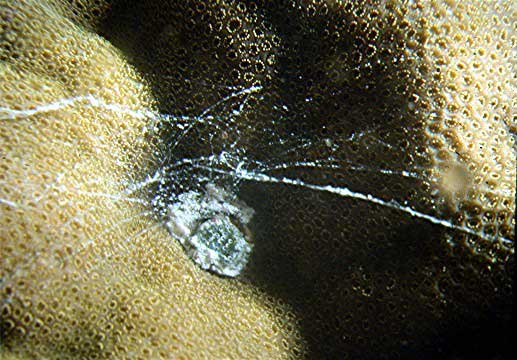
Figure 5. The feeding strands of a vermetid embedded
in a coral in Yap. If vermetids
become abundant in a reef tank, the copious production of
mucus strands may
irritate some corals. Generally, however, they are harmless.
Although I have concentrated on the worm
snails of the family Vermetidae, two other families can have
shells with a similar appearance. I consider it rather unlikely
that specimens of either of these two families would appear
in marine aquaria; however, for completeness, they are included
here. These are the Family
Siliquaridae and the Family Turritellidae (Abbott, 1974;
Abbott and Dance, 1982). Siliquarids
look quite like vermetids; however, the shell has a slit running
along its entire length. Tenagodus species can sometimes be
found embedded in sponges. Most
turritellids have a rather normal-looking coiled snail
shell with a high spire. The oddities, in the genus Vermicularia,
look like normal turritellids initially, but then uncoil and
look rather like vermetids. They may be found embedded in
colonial ascidiaceans or sponges. Some species grow attached
to gorgonians as well. Little is known about these two types
of worm snails, either about their natural history in general,
or their feeding habits, in particular.
Conclusion:
In
many reef tanks, some of the most abundant animals are these
small snails that often appear to be calcareous tube worms.
The larger species are rather rare in aquaria, but the abundance
of the larger vermetids on some Indo-Pacific reef flats is
truly striking, and gives an indication of the amount of the
appropriate detrital or particulate food available. Similarly
well-adapted for reef aquarium life, the smaller species are
sometimes prolific to the point of being nuisances. However,
in most tanks, they simply remain an example of a small, but
highly successful, component of reef biodiversity.
|