|
It has been a few months since I have written
a column. I have many ongoing projects that should make great
reading in the near future, but none of them is completed
yet, and I could not find it in me to write something about
corals just for the sake of writing. This month, I choose
to share briefly some of the more interesting findings I have
come across during the work on some of these projects. As
it turns out, the subjects all involve organisms that are
very small.
Parasitic Copepods
I have recently taken a keener interest
in the small red crustaceans that are associated with Acropora
and which have been the subject of much discussion over the
past few years. During my literary investigations into these
creatures, I came across a mass of papers by Arthur Grover
Humes, a copepod researcher who died in 1999 after having
named some 700 new species, 140 new genera and 16 new families
of copepods. His early work involved copepods that were parasitic
in various marine organisms, but beginning in the 1960s, his
fieldwork took him to Madagascar where he discovered parasitic
copepods in corals. Over a period of some twenty years, he
continued his work with corals in Mauritius, Australia, New
Caledonia, the Moluccas, Enewatok Atoll, Panama, the Philippines,
and other Indo-Pacific and Caribbean locations.
What is most remarkable about his work
is that in reading his review publications that sometimes
exceeded 50 pages each, it became apparent that virtually
every coral he examined had copepods associated with it, many
of them parasitic and many only associated with a single species
or genus of coral. Unfortunately, as a taxonomist, he was
interested more in the relationships between the copepod species
than in the natural history or ecological information about
the crustaceans, or how they might be parasitic. His remarks
about their relationships with host corals were based on anatomical
structures that are identified as being correlated with a
parasitic lifestyle. For the majority of the species he described,
no further work has been done to discover more of the nature
of these copepods. Also interesting is how they were extracted.
Humes mentions repeatedly that if one were to simply fix coral
tissues with formalin, or try to dissect its polyps, the copepods
would be missed, for they leave the polyp upon disturbance.
It was only through careful and slow introduction of graded
series of ethanol concentrations that he was able to find
the hundreds of crustaceans living within the coelenteron
of coral polyps or, occasionally, on their surfaces.
Scleractinian corals have more copepod
associates than any other cnidarians, with species belonging
to the families Anchimolgidae, Rhynchomolgidae and Xarifiidae.
Some of these are bizarre looking creatures. The Anchimolgidae
are exclusively associated with scleractinian corals and include
at least 84 species in 28 genera. The Xarifiidae are internal
parasites of both hermatypic and ahermatypic scleractinian
corals (though absent in the Caribbean), consisting of another
84 species in four genera. Within the Octocorallia, the Alcyonaceans
(soft corals) have the greatest number of copepod associates,
and 98 copepod species are now known to occur on Indo-Pacific
Alcyonaceans.
Reading this massive amount of literature
on copepods made me realize two things. First, if parasitic
copepods are so common in corals, what are their effects?
I would suspect that no small amount of stress or mortality
might occur as a result of these little crustaceans under
less than ideal conditions. Second, I also learned that the
red crustacean associated with Acroporids does not appear
to be any of the copepods that Humes described as Tegastes
and other copepods associated with Acroporids apparently inhabit
the gastric cavity. I am busy at work trying to learn more
about this troublesome red bug, and have enlisted the services
of several specialists to help with the description of the
animal, and I will write more about them in a future article
as I learn more.
This crustacean has become the bane of many aquarists keeping
certain species of Acropora.
Marine Actinomycetes
While working on the Elegance Coral Project,
I found several papers describing the presence of a marine
group of microbes within the Actinomycetes, previously well-known
only from terrestrial environments. These are aerobic, gram-positive
bacteria that form branching filaments or hyphae, produce
asexual spores, and can even produce fruiting bodies. In many
ways, they are very similar to fungi. Actinomycetes are widely
distributed in terrestrial soil and are among the most important
components of the decomposition pathways for organic matter.
They are what give an "earthy smell" to soil. Many
live in symbioses with plants, distributed near roots and
rhizomes, and providing nutrients to the plants. Actinomycetes
are also notable for another reason; they are one of the most
important producers of medically useful antibiotics. The antibiotic,
Actinomycin, for example, was one of the first antineoplastic
(anti-cancer) drugs developed from Streptomyces. It
is quite toxic, and besides being used in chemotherapy, is
an important tool in cell biology research. I also use it
in my studies on coral disease
Until recently, it was assumed that the
Actinomycetes found in marine sediments were simply saline
tolerant spores that had been washed into the ocean from land
based sources. It is now known, however, that exclusively
marine Actinomycetes are diverse and widespread. Because of
the newness of this recent discovery, little is known of the
biological and ecological roles of these fungi-like bacteria
on coral reefs, but it is likely that they play a similar
role in decomposition pathways. The antibiotic production,
symbiotic nature, and decomposition aspects of Actinomycetes
have interesting implications for tanks employing deep, fine
sand beds. I am currently working with two marine microbiologists
who specialize in Actinomycetes to determine if they have
a role in the pathology of the Elegance coral, Catalaphyllia
jardinei.
Actinomycetes are filamentous bacteria found in sediments
that share many characteristics with fungi.
Brown Jelly Bugs
The coral malady, black band disease, consists
of a consortium of somewhat variable cyanobacteria, sulfur-oxidizing
and sulfur-reducing bacteria, and a mix of other microbes
that form a coordinated self-sustaining band that consumes
coral tissue as it moves across the corallum by the actions
of sliding filaments of the cyanobacteria. The condition called
"Brown Jelly" in aquarium corals appears to share
many similar traits, albeit a very different appearance. I
have run into a wall with regards to my investigation of brown
jelly in corals. One of the difficulties of studying this
condition is that it is uncommon, and consequently, it is
not easy to get samples or conduct experiments. Like black
band disease, brown jelly has a rather distinctive appearance
and is composed of a diverse assemblage of microbes. What
is still unclear is whether any of the microbial community
causes tissue death, or if the tissue is already dying and
the jelly is consuming dead tissue. The jelly can act as a
contagious agent, and corals that contact the jelly may subsequently
die with the presence of increasing amounts of brown jelly.
This material could, however, simply be smothering the coral
tissue, while consisting of a group of self-sustaining and
rapidly multiplying constituents that do not actually constitute
an "infection."
As I have mentioned before, the ciliates
that have been presumed to "cause" brown jelly do
not appear to cause it at all. They are consuming zooxanthellae
and other material released from dying and decomposing coral
tissue - performing a janitorial role of sorts. In analyzing
the material, it is complex in its composition, including
many microbes which I am wholly incapable of identifying.
I asked a coral disease colleague of mine, Debbie Santavy,
if she knew anyone who is a good protozoologist. As it turned
out, her husband Richard Snyder, happened to be one. I left
some brown jelly with him, and recently received his preliminary
analysis.
He found the flocculent material to be
dominated by algal cells, most of which were Symbiodinium
(zooxanthellae). There was a fungal component that provided
some structure to the material. He found several sizes and
shapes of fecal pellets that contributed to the brown color
also provided by the golden brown zooxanthellae and suspected
they originated from copepods. I had noticed many Spirochaetes
in the mix, which he identified as Spirulina. He also
found flagellates, which were likely the very small fast-moving
bugs I saw associated with the dying tissue. He suspected
there would be a lot of amoebae present in the sample given
the habitat, but they were not identified in the mix because
they are easily lost in the fixation process. In contrast
to my inexperienced eye noticing a couple types of ciliates,
he was able to identify five or six different species, although
none could be positively identified to a specific species.
One of these ciliates was the species feeding on the zooxanthellae.
There were also abundant bacteriovores feeding on the even
more abundant heterotrophic bacteria, composed mostly of filaments
and rods.
He suspected the microbial activity in
the production and binding of polysaccharides was responsible
for the material's gel-like flocculent consistency. In closing,
he noted that he did not notice anything, except perhaps the
fungi, that would cause harm to the corals. To quote, "I
would consider this consortium to be your friend, flocculating
the extraneous cast-off organisms and organic material so
that you can keep it siphoned out of your tanks." That
would be true were this material not able to cause further
coral mortality as it grows and is blown around the tank by
water currents. It is clear that there needs to be a lot of
work done in isolating and culturing each of the consortium
members to determine if any one of them actually causes tissue
death in corals. In fact, it would require a considerable
amount of time, effort, and money. I plan to injure some corals
and keep them in rather stagnant tank water to see if I can
induce a "brown jelly infection" and send Richard
some live material. If I can find a way to reliably and consistently
produce or culture brown jelly, it would make some sense to
continue this work. With available material, I could try to
infect corals by selectively treating the jelly with drugs
that could eliminate some groups of the consortium and determine
their individual effects. Otherwise, it appears that brown
jelly will remain a mystery for the time being.
 |
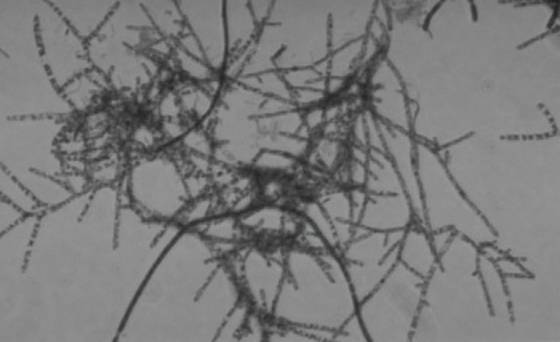
This Hydnophora sp. is affected with brown jelly, a
condition well known to aquarists. The cause
of coral mortality
by this material remains unknown.
An Unrelated Story That May Be Of Some Interest…
While not involving microscopic organisms,
I have one final anecdotal report to provide to readers of
this column. For nine years, I have had a soft coral in my
tanks that has been spread extensively around the country.
I originally acquired it from Scientific Corals, in Atlanta,
as Litophyton arboreum. I am not convinced it was identified
correctly, and it appears to me from examining its sclerites
and gross colony and polyp morphology to be a species of Capnella.
In any event, I have this colony spread throughout five tanks
in my home and lab. It is a prolific producer of daughter
colonies by branchlet dropping. It is also worth noting that
both Capnella and Litophyton have very high
densities of zooxanthellae, and are considered quite close
to autotrophic corals, at least in terms of carbon produced
by photosynthesis. Thus, one would expect their growth to
be strongly correlated with irradiance levels.
Over the past three months, I have provided
46 daughter colonies to a local coral farm. What made me think
about this was that almost all of the colonies were being
produced from parent colonies in my large home system. I began
thinking about the different tanks, and the relative contributions
of light and food to growth and reproduction. While the tanks
are, of course, very different they all have rock and sand
that has been intermixed between them many times. These tanks
all have seawater made from the same salts, and I use the
same regimen of additions and maintenance on all of them,
including food, calcium and alkalinity and nothing else. Water
changes have not occurred at all during the period in question.
The water quality parameters I measure are nearly identical
and their values certainly fall within the variation found
on natural reefs. Thus, I would consider them to be different,
but not so different as to be somewhat like the differences
between various reefs where a species is known to exist.
The tanks are described in the table below,
and the time period is approximately three months to date.
| |
Tank size
(gallons)
|
Light
|
Skimmer
|
Colony sizes
|
Colony growth
|
# Daughters
|
| Tank
1 |
10
|
2 x 65W power compact
(one blue, one white)
|
None
|
Adult - 10cm max
|
Rapid
|
4
|
| Tank
2 |
10
|
18W fluorescent
white actinic
|
None
|
Sub-adult, 3 cm
|
Imperceptible
|
0
|
| Tank
3 |
75
|
2 x 65W power compact
(one blue, one white)
|
Tunze - estimated
low efficiency
|
Sub-adult, 5 cm
|
Slow
|
0
|
| Tank
4 |
120
|
2 x 400W 6500K
|
My Reef Creations,
Beckett design, estimated high efficiency
|
Adult, 10cm max
|
Rapid
|
46
|
| Tank
5 |
55
|
2 x 175W 10,000K
|
None
|
Sub-adult, 5 cm
|
Slow to moderate
|
0
|
The variations in the number of daughter
colonies produced, and the growth of colonies in the tank,
is remarkably different under the different conditions. While
many other variables might be involved, such as water motion,
competition, and differences in food source amounts and types
produced in situ from the various tanks, it is most
likely that growth and reproduction are most directly attributable
to energy availability. The difference in placement of the
corals with respect to depth and subsequent light attenuation
is assumed to be minimal since multiple colonies in each tank
are found within similar ranges in all tanks.
So, one might assume that Tank 2 is simply
too low in irradiance, and that the skimmer in Tank 3 is removing
food sources that are present in tank 1, despite identical
irradiance. Tank 1 produces daughters and adult colonies,
while Tank 3 produces neither. In Tank 4, the very high light
level might be responsible for the daughter colonies and rapid
growth, but Tank 5 is an anomaly. It has higher irradiance
than Tank 1; neither tank provides export of any kind, yet
Tank 5 produces no daughter colonies or adult colonies.
Of course, none of this really means much.
This is simply an anecdotal tidbit that I found interesting,
and I offer it because it should at the very least spark some
more debate in the Coral Forum with regard to light versus
food in coral growth. It has become the aquarist's equivalent
of heredity versus environment, and sometimes having so many
tanks running with the same clonal lines gives me an opportunity
to throw things like this out to the masses, if only for fun
and lively discussion.
|