|
One of the more fun things that invertebrate
zoologists do is to speculate on the relationships of the
various groups of animals. One question, "Who evolved
from whom?" has dominated and driven research about these
animals for a very long time. Until recently many of the discussions
about evolutionary relationships have been exercises in logic
based on limited evidence. This has led to a lot of interesting
and, generally, fun discussions about which character sets
were most important in determining relationships (the important
ones were always the ones "you" chose) and which
weren't (always the other guy's). In essence, as we had no
real way of independently evaluating the various lines of
evidence and conjecture, such discussions were not really
science, but more akin to science fiction.
Recently, however, with the advent of techniques
allowing for direct comparisons of the genetic code of organisms,
it has become possible to actually assess the degrees of genetic
interrelationship amongst and between the various major animal
groups. Additionally, the development of a particular set
of statistical techniques and methodologies called "phylogenetic
systematics" has allowed a separate independent way of
assessing relationships based on evolved or derived characters
shared between animal groups. In essence, we now have some
tools that permit us to get answers to the same question from
two independent methodologies. Consequently, we can use the
two methodologies together to check on each other and to assure
ourselves that the results are no longer in the realm of literature,
but rather actually are "real" science.
Over the past ten to fifteen years, these
two tools have allowed us to put together a relational framework
of the animal kingdom that has overturned some long-cherished
and well-defended ideas. These were ideas that seemed logical
at the time, but were simply wrong; and in hindsight, one
wonders how they could have been supported for so long. One
of these cherished ideas held that the segmented annelid worms
and the segmented arthropods were closely related simply because
they both were segmented and had an internal morphology that
was fundamentally different from the third great segmented
group, the vertebrates.
One of the more striking discoveries of
the new methodologies was that there are three main lineages
or assemblages of animal groups. One lineage, called the deuterostomes,
derives the Echinoderms (sea stars, sea urchins, and related
animals) and Chordates (fishes, mammals, and birds) from the
Cnidaria (corals, sea anemones and hydroids). This linkage
was not too dissimilar to what had been proposed prior to
the new methodologies and served, in some regards, to validate
the logical methodology used through most of the nineteenth
and twentieth centuries. However, the lineages presumed to
link the rest of the animal kingdom, indeed the vast majority
of the animal groups, was thoroughly juggled and rearranged.
We now recognize one vast assemblage of many animal groups
which is defined by the presence of a particular type of larva
called a trochophore, or the presence of a crown of ciliated
tentacles around their mouth. This crown of tentacles is called
a "lophophore." This assemblage of animal groups,
containing the mollusks, annelids, brachiopods, and numerous
smaller groups is called the "Lophotrochozoa," a
name derived from the names of the two major characters, the
lophophore and the trochophore.
The final assemblage of animal groups,
containing relatively few major groups but, by far and away,
most of the animal species, is characterized by animals that
have to molt to grow. These animals have either an inelastic
cuticle or an "exoskeleton" that covers their entire
body. The name for the group, the Ecdysozoa reflects their
need to shed their outer covering as they grow. Ecdysozoa
is derived from the words "ec-" (from the
Greek word "ek-" meaning: "out of"), +
"dysis" (from the Greek word, "dyō,"
meaning: "to put on clothes,") giving the New Latin
word, "ecdysis," (meaning: "a getting
out of clothing, molting.") +"zoa" (from
the Greek word, "zōon," meaning: an animal
or living being). So, these are the animals that molt or get
out of their clothes or, in other words, they are strippers.
The Strippers…
Apparently the genetic machinery for the
construction of a cuticle or inelastic covering, coupled with
the need to shed such a covering to grow really evolved only
once as all of these groups seem to have the same basic genome
for the physiological "machinery" necessary for
molting. There is a progression of forms from simple molted
cuticles to quite complex molted "exoskeletons."
The latter, of course, are found in the arthropods, such as
crabs, insects, and spiders, whereas the simpler forms are
found in several other animal groups or phyla. Complex, but
simpler, cuticles are also found in nematodes or roundworms.
Both of these groups are quite well represented in aquaria.
I will discuss nematodes and the arthropodan
groups in some subsequent columns, but in this column I thought
it would be fun to explore some of the lesser-known strippers.
Molted cuticles are found on animals in many groups that are
actually reasonably common on coral reefs, and it is likely
that some of these animals have found their way into reef
aquaria, even though aquarists may not have yet noticed them.
Subsequent to discussing odd groups in this column, I have
often been asked questions about representatives that have
been recognized as living in someone's aquarium. Perhaps that
will be the case here.
With only a few exceptions, these lesser-known
animals are really quite small, and this contributes to the
lack of information about them in the hobby. There are three
groups of animals that I will be discussing in this column,
the Priapulids, the Kinorhynchs, and the Loriciferans. Only
the Priapulids are likely large enough to be commonly visible
to aquarists. The Kinorhynchs would certainly be visible to
those aquarists who periodically microscopically examine some
of their systems' sediments. Loriciferates may or may not
be present in aquaria; they should be there, but are rarely
seen anywhere.
Priapulids
Priapulids are some rather odd-looking
worms and take their common name from their taxonomic placement
in the phylum Priapulida. These are generally small animals
which, although they may appear annulated, do not have a body
comprised of separate segments. There are about 20 extant
species, and they are found in all oceans, although they are
most diverse in coral reef areas. There are no fresh water
species.
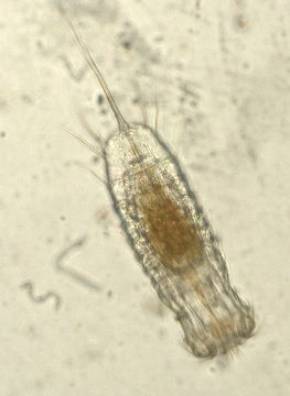
Figure 1. Priapulus caudatus, a widely distributed
Priapulid worm.
The largest of them reach lengths of about
four inches. They have three body regions. In the front is
a spiny introvert or proboscis, which has occasionally been
dignified with the name of "head," however these
animals really don't have a defined head. This structure is
covered with spines which are used to grab the prey and possibly
also to aid in burrowing. The main part of the body is called
the trunk, and it is annulated; that is, it appears to be
comprised of rings of tissue. Off the back end of the worm
are generally one or more structures referred to as caudal
appendages. These may serve a respiratory function.
Here is a link to a diagram showing several
species of Priapulids:
http://eebweb.arizona.edu/globe/priapula.jpg
The body is completely covered with a cuticle,
and as is characteristic of all of the animals discussed this
month, this cuticle needs to be molted to allow the animal
to grow. The cuticle is made of protein when the animal is
young, but becomes chitinized as the animal grows. The polymer,
chitin, is found in many invertebrate cuticles. It a very
good structural material as it is very durable and chemically
resistant, yet it is flexible enough to allow a wide variety
of movements.
Here is a link to information about a Priapulid,
Tubiluchus, that is different from the commonly illustrated
Priapulus:
http://www.ldeo.columbia.edu/dees/ees/life/slides/phyla/priapulida.html
Priapulids have a simple nervous system,
consisting of a simple ring around the esophagus, and a midventral
nerve cord. There is no really well-defined brain, and little
is known about their nervous system. The body wall is generally
quite tough and muscular, and the muscles are striated, indicating
that the animals are capable of quite rapid movements.
Little is known about their natural history,
but they appear to live in burrows and perhaps strike out
of their burrow to grab prey, which they seize and engulf
with their proboscis. Some species may live head down in burrows
foraging in sediments or in rocky crevices for prey. It appears
that most coral reef species are found in the reef rocks or
reef rubble, and these are the Priapulids which may be expected
to appear in aquaria from time to time.
Figure 2. The mouth of Priapulus caudatus. Note
the very "functional" teeth used to hang on to prey.
Quite surprisingly for a group of wholly
soft-bodied animals, Priapulids actually have a significant
fossil record. Although they are absent from large spans of
the Earth's fossil history, they are prominent and dominant
animals in some of the earliest good deposits of animal fossils,
such as the Burgess Shale Fauna, and similar faunas from southeastern
China. There are 11 species of presumed Priapulids found in
the Burgess Shales, a fossil deposit dating from about 530
million years ago. Interestingly enough, Priapulids are more
diverse in this deposit than they are in any present day marine
habitat.
Here are some links to the Burgess Shale
Priapulids especially Ottoia:
http://astrobiology.ucla.edu/ESS116/L02/0254%20Burgess%20worm.jpg
http://park.org/Canada/Museum/burgessshale/worms.html
http://www.dc.peachnet.edu/~pgore/geology/geo102/burgess/priap.gif
Kinorhynchs
The next group of small strippers, is the
Phylum Kinorhyncha; or as they are know to everybody who has
seen one, the kinorhynchs. The name of this small group is
derived from the Greek words, kineō meaning moving
and rhynchos meaning a beak or snout. The name is very
apt as the animals are continually moving and feeding by pushing
their introvertable proboscis in and out. There are only about
one hundred species of scientifically described kinorhynchs,
although most experts seem to agree that the total species
number likely significantly exceeds that rather paltry sum.
Figure 3. Two different species of Kinorhynchs. Left
image. View from
above the animal. Right image: Lateral view.
All kinorhynchs live in marine habitats,
and they are all small, generally less than about 1/25th of
an inch, or 1 millimeter in length. They live in sand or sandy
mud and are not uncommon in such environments throughout the
world. They likely enter reef aquaria within wild collected
live sands. Although, they have not been definitively reported
from reef aquaria, I would expect them to be present in some
tanks, and they may actually be quite common, if overlooked,
reef tank inhabitants.
Here is a link to a labeled diagram and
some information:
http://www.ldeo.columbia.edu/dees/ees/life/slides/phyla/kinorhyncha.html
And another link showing some of the external
features:
http://biodidac.bio.uottawa.ca/Thumbnails/showimage.cfm?File_name=KINO001B&File_type=GIF
These are segmented animals, and generally
have thirteen segments, referred to a zonites. The body and
segments are often covered by spines, but they lack articulating
appendages. The most anterior point of the animal is the mouth,
located on an introvertable cone, which can be withdrawn into
the body. The body is covered with a thick chitinous cuticle.
There are three chitinous plates that comprise each zonite,
one thick curved dorsal plate, and two thinner ventral plates.
Depending upon the species, there are often adhesive tubules
that open in the ventral region of the worm. These are used
to "fasten" the animal in place when it is feeding.
Figure 4. Lateral view of a Kinorhynch.
Kinorhynchs have a number of interesting
attributes, not the least of which is the fact that the epidermis
underlying the cuticle is a syncytium. That is to say, it
lacks cells, and basically is a mass of protoplasm with many
nuclei in it, but no cell membranes. This layer overlies a
highly muscular dermis with striated muscles. These are rapidly
moving little worms and the relatively complicate musculature
that furnishes that movement is comprised of longitudinal,
oblique and dorsoventral muscles. The nervous system is relatively
complicated, and built in the invertebrate pattern with a
ring of nervous tissue surrounding the mouth. There are a
wide variety of sensory structures, such as sensory spines
called scalids, as well as ocelli, or eyespots, in some species.
The gut is complex and with a highly muscular
throat region where the cuticle is often very thick. Kinorhynchs
are often described as grazers on diatoms and other sediment
dwelling algae. The muscular, cuticularized throat region
likely acts as a gizzard, crushing the prey for subsequent
easy digestion.
Kinorhynchs have separate sexes, and internal
fertilization, although copulation has not been documented
in the scientific literature. The female lays eggs which hatch
to give a juvenile of eleven segments. These animals grow
by molting and with each molt a segment is added until they
reach the adult complement of thirteen segments. Any subsequent
molts result in an increase in size, but no additional segments
are added.
These small worms are likely reasonably
common in some reef aquaria, but presently are unknown in
reef tanks. Because of their small size, it will likely be
necessary to examine the sediments microscopically to find
them. If they are found in reef aquaria, they will likely
contribute to the "clean up" crew, eating diatoms
and moving through the sediments, thus facilitating the functionality
of the sediment biological filter.
Loriciferans
The last group discussed in this column
will be the Phylum Loricifera. As with the other two phyla
discussed this month, they have no common name and are referred
to as Loriciferans. The name refers to the "Lorica"
or "shelled" external body wall that they possess.
They are also unlikely to be observed in aquaria; but who
knows, perhaps they will be seen and recognized in some lucky
soul's system.
As I have no images of my own for the Loriciferans,
you should follow these links to find some nice images of
them:
http://www.meiofauna-marina.com/contents/images/loricifera.jpg
http://whyfiles.org/022critters/images/lori3.gif
And this link will lead you to some labeled
diagrams, so that you might appreciate just how strange are
the structures in these animals:
http://www.ldeo.columbia.edu/dees/ees/life/slides/phyla/loricifera.html
Loriciferans were described as a phylum
in 1983, although the animals were first seen about 20 years
earlier. These are animals that are best described as bizarre,
and yet they are structurally amongst the "neatest"
looking invertebrates. They look rather like miniature mechanical
constructions rather than as members of the animal kingdom.
Only about a dozen species are known, and none have been seen
alive for more than a few minutes. They are minute, and may
be the smallest of metazoan animals.
The body is regionated, and as with both
the Priapulids and the Kinorhynchs, they have an introvertable,
quite spiny, head. There is also a large abdomen, enclosed
in a cuticular shell or lorica consisting of several rows
of plates and structures arranged in a pattern of sixes. The
mouth is located at the tip of the "head cone" much
as in the kinorhynchs, and there are protrusible stylets or
spines in the cone. The throat region contains what appears
to be a muscular pumping apparatus, and this region is also
lined with a molted cuticle. Although the diet is unknown,
it appears that the throat area could act as a gizzard to
crush or mash any ingested material.
The nervous system is relatively quite
large, and as in the Kinorhynchs and Arthropods, virtually
every spine on the body appears to be enervated by sensory
nerve fibers. These animals receive a lot of sensory input
through their cuticle by means of the sensory spines.
Remarkably little is known about Loriciferans,
even though they have been studied for about thirty years.
They glue themselves to sediments and are commonly found in
relatively coarse shell fragment sands near Floridian and
other reefs, but the only way to get them out of the sediments
is by shocking the sediments with fresh water. As many aquarists
realize, such fresh water rinses kill most animals, and that
is also the case with Loriciferans. However, such rinses also
break them free of the sediments, and their corpses can be
collect for examination and study. It is hard to do natural
historical or ecological studies from dead animals, and this
is compounded by the fact that these are exceptionally small
animals, so it is particularly difficult to identify gut contents.
Their presence around Floridian, and other reefs, argues for
the likelihood of their presence in reef aquaria, but obviously
they have never been documented from aquaria.
Loriciferans will only be noticed if an
aquarist undertakes a particular search for them. Sediments
will have to be removed from the aquarium, and rapidly rinsed
in fresh water to remove adherent materials and animals. The
material cleaned out of the sediments by the fresh water rinse
has to be rapidly sieved through a fine screen, preserved,
and then microscopically examined. As they seem to be relatively
rare and they are very tiny, it will likely take a lot of
looking before any are seen. Of course, if any are seen, it
would be momentous enough to warrant publication in both the
scientific and popular literature. If anybody participates
in a Loriciferate safari, I heartily extend my good wishes
for good luck! These are oddest of animals and it would be
grand to find them in aquaria.
|