A
Propensity to Interfere and the "Geology" of Coral
Reef Aquariums
Over the years, I
have moderated or participated in many coral and aquarium
related web sites, message boards, and mailing lists. One
of the reasons I do so is that it allows me to keep a "pulse"
on the aquarium hobby; getting an overall feel for the status
of both the relative pervasiveness of information and techniques
throughout the hobby, as well as knowing, perhaps in advance,
when certain trends are beginning to emerge.
When I began moderating
the AOL reef aquarium message boards many years ago, certain
facets of aquarium and coral husbandry were obvious and
trends came and went over the years. Some stayed, some are
long forgotten or, for newcomers, were never known: the
widespread use of wet/dry filters, the use of novel additives
(Molybdenum, Vitamin C, Liquid Gold, Boron supplements,
Coral Vital, sugar, various "X-products", yeast,
Lugol's solution, etc.), plenums, sulfur lights, blacklights,
and so forth.
|
|
|
An advertisement
for a "miracle product"
1992. Freshwater and Marine Aquarium 15 (4): 174.
|
This first article for Reef
Central's exciting new online magazine is the beginning
of my plan for addressing some of the more prominent themes
that emerge from The Coral Forum board. I would like to
take some of the more common "threads" and look
at them in some depth; essentially offering not only more
information about some of those things that seem to be causing
either questions or troubles for many aquarists, but perhaps
efficiently addressing some far reaching areas of misunderstanding.
As usual, I will be drawing my information from my experiences
in the field and with aquariums, as well as from scientific
literature and, where possible and useful, aquarium observations
and anecdote.
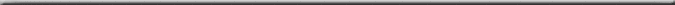
Part One:
The Coral Reef Aquarium: Operating Room, Emergency
Room, or both?
One thing that has certainly
not changed over the years is our seemingly irrepressible
need to intervene into the normal function of an aquarium.
It has been known for quite some time that it is usually
better to keep one's hands out of the tank, yet this does
not seem possible for most. The topic for this issue was
overwhelmingly obvious to me, and had caused me several
hours of private consternation. One phrase typed many times
in The Coral Forum over the past several months keeps coming
back to haunt my thoughts:
"Should I cut it out?"
Why is it that aquarists
seem to be budding surgeons? Is it perhaps that we are so
attached to our corals that we tend to anthropomorphize,
pretending as if they were patients of ours, requiring constant
medical supervision? Is it the fear of the unknown pathogen?
It is a concept that has so firmly entrenched itself in
the minds of aquarists around the world that it is frightening.
Coral bleaching, coral disease, coral predators. What is
it? Is it harmful? Will it kill my coral? Will it spread?
DO I NEED
TO CUT IT OUT??!!
I don't want to poke fun
at those genuinely concerned and empathetic aquarists who
are so caringly worried for their animals. In fact, it's
great that you all have such a strong attachment that you
would be willing to brave the waters of unknown pathogens,
predators and diseases with your bare hands to rescue this
poor suffering cnidarian!
So, is the aquarium an operating
room? Yes, occasionally. There are indeed times when the
natural regenerative function of these colonial animals
can be put to great advantage. Excision can be done, effectively
and with minimal risk, in many cases. But when are those
cases? Soon, I will attempt to answer that question.
Is the aquarium an emergency
room where corals appear mortally wounded with sometimes-slipshod
procedures performed in order to stabilize a condition that
requires long-term treatment? Is it a place where corals
are admitted when it's a false alarm, like the patient who
ate too many jalepenos and thought that pain in his abdomen
was an abdominal aortic aneurysm when all he needed was
a Maalox? The answer is a resounding yes.
The question that
begs to be answered is "What's wrong and just how serious
is it?" The question is rarely, "Do I need to
cut it out?"
Part Two:
Some Facts About Colonial Animals
A colonial animal
is one where groups of related organisms live together,
sometimes interconnected and even indistinguishable from
one another. We have discussed aspects of this subject in
other threads in The
Coral Forum. In the case of colonial organisms like
many of the stony corals, groups of polyps live in various
degrees of connection with others, sharing the same skeleton,
but each polyp residing in its own skeletal element called
a corallite. In some, like the genus Acropora, the
polyps are highly interconnected and share tissue elements,
such as gastrodermal canals and nerve net elements. Even
in genera of corals with somewhat less connection, the tissue
is often unified across the skeletal surface (corallum)
and this tissue is termed to coenosteum. There are stony
corals with even less connection that are still considered
colonial. For example, Caulastrea spp. and some Euphyllia
spp., often reside within the same major skeletal elements,
but each polyp is separate from all the others, having divided
from a parent by fission and then continuing off in its
own direction, calcifying more of its own corallite separately
from its parents, until it too divides. Soft corals, corallimorphs
and zoanthids are similar; some have polyps inextricably
fused in a mass of tissue called the mesoglea. Others are
almost totally separate, perhaps connected by a thin sheet
or mat, or not at all.
|
|
| This Euphyllia parancora
is an example of a coral with separate polyps that still
forms colonies. The individual polyps become disconnected
from each other over time, although the newly dividing
polyp in the foreground is still connected to each half.
Here, one can also see the dead parts of the skeleton,
and dead polyps. These fragments form the base of a
low relief reef comprised entirely of broken corals
and are the substrate for fields of Goniopora
spp. and Nemenzophyllia turbida. Eventually,
the amounts of bioerosion, already clearly visible in
the upper right, will become sand. Right now, it is
home for sponges, tunicates, and other encrusting and
boring life. |
So what does this mean?
Well, in the case of the highly interconnected types, what
affects one polyp may well affect numerous polyps. This
spans the range from nutrition to local necrosis. With the
individual polyps, if a polyp dies, unless the other polyps
are subjected to the same effector, it dies alone. In a
highly interconnected colony, it's more difficult to separate
the influence and prevent it from affecting other polyps.
However, such colonies have other aspects that have given
them an evolutionary edge. In living in large accumulations
of numerous individuals, a colony is often able to tolerate
some degree of mortality or damage without the loss affecting
the entire colony. In fact, a coral reef is such a highly
competitive environment, this type of advantage is almost
mandatory to ensure the continued reproductive presence
of such important components of the ecosystem. As such,
coral colonies are extremely long lived and show limited
senescence and yet are almost always enduring alternating
periods of growth and set-back. The term used to describe
this is partial mortality.
|
|
| Two species of coral, a Porites
sp. and an Oxypora sp., battle it out on the
reef. The dead zone between them is covered with cyanobacteria,
itself an important component of coral reefs. This dead
space can be settled by a new variety of life, or the
dominant coral can continue to grow. The Porites
sp., a competitive subordinate, has the advanatge in
that it tolerates partial moratality with ease and is
extremely long lived. |
The Geology
and Growth of Corals, Coral Reefs, and Coral Reef Aquariums.
Corals grow by precipitating
calcium carbonate underneath their lower polyp surface using
cells of the calicoblastic epithelium. This is the same
tissue layer that forms the upper surface of the coral that
is visible, but the cells are slightly different and attached
to the skeleton itself. Over a continuous time frame that
can span centuries, huge formations of calcium carbonate
are formed that, along with the biomineralizations of crustose
red algae (coralline algae), foraminiferans, mollusks, bryozoans,
and other calcifying reef biota, comprise coral reefs. The
carbonate skeletons of these animals, long dead, provide
for the complexity of surfaces and small niche habitats
that allow for the huge biodiversity of life on a coral
reef. Animals and plants live around these structures, within
the cracks and crevices used as both home and shelter, and
even within the pore structure of the carbonate itself.
Boring organisms make their home inside the solid skeletons,
and many such eroding organisms are ultimately responsible
for the production of dissolved calcium carbonate in seawater
that, in turn, feeds more calcification. Furthermore, old
skeletons are ground up by mechanical, chemical and biological
processes to form the sands that support their own integral
communities; from decomposition areas to plain benthic habitats
for an abundance of lifeforms. As coral reefs grow, they
gain surface complexity and area, and are thus able to support
even more life.
 |
| This Leptoria sp, is a colonial
group of polyps so integrated that the skeleton formed
by the polyps is even incapable of distinguishing the
fused polyps that lie side-by-side within the sinuous
meanders. |
Corals
that die only lose the thin veneer of tissue that created
the massive carbonate skeletons beneath them. This surface
then becomes an important settlement area for new plants,
animals, and even coral planulae that will continue forming
the outward growing margins of the reef. An entirely dead
coral colony merely becomes habitat and substrate for further
growth. Most of the time, barring massive mortality from
bleaching, disease, competition, predation, storm, etc.,
any mature coral colony on a reef will have a very high
likelihood of having suffered at least some partial mortality
over its potentially extremely long history since settling
as a tiny planula. In some cases, the partial mortality
may have occurred because part of the colony was growing
under suboptimal conditions. Perhaps some branches of a
colony had grown too close to a strong competitor, or had
grown under a shaded ledge and could not receive adequate
light. In this case, the partial mortality might even be
considered adaptive as the colony can expend energy in growing
outwards in more favorable areas.
In fact, ideal coral
reef growth is considered to be accomplished under what
is termed the "periodic disturbance hypothesis."
That is, coral reefs maintain their highest diversity and
health when periodically subjected to disturbances that
cause partial mortality. This prevents any of the fiercely
competitive plants and animals from achieving spatial and
ecological dominance. It"clears the brush," so
to speak, much as forest fires renew forests over the long
term, and allows for the introduction, settlement, and growth
of new genetic and ecological diversity that ultimately
benefits the reef as a whole.
|
|
| This solitary polyped coral, Scolymia
sp., cannot tolerate partial mortality. It can, however,
tolerate injury even to the extent that skeleton is
exposed. |
In
aquariums, we have a vested interest in the life behind
the glass. Hopefully, it is more than just a financial interest,
although this may be a strong motivator, as well. We try
very hard to separate each individual "specimen,"
protecting it from the ravages of competition. We place
it carefully as a tiny juvenile colony to maximize its health,
and then, as our efforts succeed and the coral grows, we
decide to play doctor and landscaper, trying to maintain
a miniature dollhouse of corals.
|
•
|
If a coral starts
to grow large, what do we do? We cut it out. |
|
•
|
If a coral grows
too close to another, what do we do? We cut it out. |
|
•
|
If a coral suffers
some partial mortality and results in something temporarily
less attractive, what do we do? We cut it out. |
|
•
|
Disease? Cut it
out. |
|
•
|
Bleaching? Cut
it out. |
|
•
|
Propagation? Cut
it out. |
|
•
|
Came in with a
small dead spot? Cut it out. |
|
•
|
This branch is
brown? Cut it out. |
|
•
|
Need room for
a new specimen? Well, you know how to make room by now!
You just cut it out. |
What's Wrong With This Picture?
Almost too much to consider.
Can you imagine what would happen to a coral reef if everytime
a death occurred, it was removed? Given the description
of coral reef growth above, it would obviously not be a
great benefit to the net growth or health of a reef. So
why do we feel it is good for our tanks?
Some might feel that is
because of aesthetics. I would argue that good money is
paid for high quality live rock, and that is exactly what
is being removed every time a dead coral is removed from
the aquarium - a future piece of high quality aquacultured
live rock. Additionally lost is net growth of a reef aquarium,
countless unseen plants and animals residing within the
skeleton, areas of potential microbial processes such as
denitrification, and spatial complexity that ultimately
makes a tank look like a reef and not a pile of rocks with
coral placed on them. When one considers it carefully, aesthetics
are lost over time, the continuous change that characterizes
a coral reef is lost, and the tank remains more or less
unchanging and stagnant from this compulsive behavior. If
coral dies in the aquarium, the only thing that really happens
is there is now an exposed surface for new settlement.
Like live rock, the
first colonizers will probably be filamentous algae. This
is normally thought of as a "bad thing to have"
in aquariums. But, if the reef aquarium is generally healthy,
this is a temporary succession that will provide some new
food for herbivores. They will graze it, grow, and reproduce.
Over time, other things will replace the algae. Each of
these successions will be important for some community in
the tank, perhaps even for the development of some species
that was latently existing in some dark crevice of the aquarium,
unseen, without an area for settlement, like a beautiful
sponge or colony of fanworms.
 |
| This Acropora sp. is dependent
on fragmentation for its ecological dominace of areas
produced by such asexual spread. The thin delicate branches
house many polyps, well interconnected, and yet the
colony is quite tolerant of breakage as a whole. |
What
else is wrong with the notion of "cutting it out?"
Corals live on the edge, delicately balanced in most cases
on a bare minimal energy budget from light and limited food
availability. In aquariums, it is probably even more of
an issue because of the limitations of a closed water volume
and the amount of food availability. As corals reach a certain
size or age, if the energy is available, they may reach
reproductive maturity and spawn. It is, to this day, an
exciting and applaudable event to have coral spawn in the
tank. If, however, a coral is subjected to constant pruning,
the energy needed to provide injury repair and reallocation
of energy to growth may compromise the coral and it may
never reproduce. In fact, with enough pruning, its growth
may be significantly slowed, as well.
Furthermore, pruning
corals results in tissue injury that compromises its integrity
and can allow for the invasion of potentially deleterious
microorganisms, perhaps even the dreaded "mystery pathogen."
It is generally found that pruning and fragmentation is
well tolerated in a healthy coral reef and coral reef aquarium,
but the fact remains that it does pose some risk to both
parent colony and produced fragment. Constant pruning also
entails the constant introduction of hands into the aquarium,
and thus increases risks of tank mishaps and the introduction
of contaminants, toxins, and non-native microbial flora.
Such non-native flora has a long history of producing problems
in coral reefs, and the implications within a closed system
are even more likely to be problematic.
Cut It
Out!
Is there ever a time
for the scissors and knives to be taken from their holders
and used? Of course. Necrotic areas that may compromise
the integrity of the entire colony should be excised. Intentional
propagation, if it is the aquarist's goal, requires some
amount of cutting or breaking. If a known disease is present
and spreading, excision is often a good treatment method.
However, a dead branch or corallite is not a reason for
excision. Competition is rarely if ever a reason for removal
of parts of a coral. Certainly, a coral that has already
died does not need to be removed - unless of course the
aquarist is someone like me who is dying to know the identification
of what species was present in the once living coral!
|
|
| I finally removed this dead Trachyphyllia
from one of my aquariums because I needed to look at
its skeleton for comparison to another skeletal morph.
The enrusting polychaete tubes are only part of the
abundant life that would have eventually turned this
coral skeleton into a small version of a coral reef
or piece of live rock. |
In summary, I would
urge aquarists to begin to think of their tanks as they
aspire them to be; living fragments of a coral reef that
are subject to all the ups and downs, the cycles, the growth
and decline; the life and the death of coral reefs. It's
probably in the best interest of your tank, and you can
keep those scissors and knives from rusting, too.
Keep reading, keep posting,
keep learning, and keep sharing.
Until then, I will
see you all in The
Coral Forum.